
疼痛生物化学:�所有疼痛综合征都有炎症特征。 炎症特征可能因人而异,也可能在不同时间在一个人身上发生变化。 疼痛综合征的治疗就是要了解这种炎症特征。 疼痛综合征通过药物、外科手术或两者兼而有之进行治疗。 目标是抑制/抑制炎症介质的产生。 成功的结果是减少炎症,当然也减少疼痛。
内容
疼痛的生化
目标:
- 谁是关键球员
- 生化机制是什么?
- 后果是什么?
炎症回顾:
主要参与者
 为什么我的肩膀会痛? 肩痛的神经解剖学和生化基础综述
为什么我的肩膀会痛? 肩痛的神经解剖学和生化基础综述
摘要
如果病人问“为什么我的肩膀会疼?”谈话会很快转向科学理论,有时甚至是未经证实的猜想。 临床医生经常意识到他们解释的科学基础的局限性,表明我们对肩痛本质的理解不完整。 本综述采用系统的方法来帮助回答与肩痛相关的基本问题,以期为未来的研究和治疗肩痛的新方法提供见解。 我们将探讨 (1) 外周感受器、(2) 外周疼痛处理或“伤害感受”、(3) 脊髓、(4) 大脑、(5) 肩部感受器的位置和 (6) 的作用。 ) 肩部的神经解剖学。 我们还考虑了这些因素如何导致肩痛的临床表现、诊断和治疗的变异性。 通过这种方式,我们旨在概述肩部疼痛中外周疼痛检测系统的组成部分和中枢疼痛处理机制,它们相互作用产生临床疼痛。
简介:对临床医生至关重要的疼痛科学简史
总的来说,疼痛的性质在过去的一个世纪里一直是一个争议很大的话题。 在 17 世纪,笛卡尔的理论 1 提出,疼痛的强度与相关组织损伤的数量直接相关,并且疼痛是通过一种不同的途径进行处理的。 许多早期的理论都依赖于这种所谓的“二元论”笛卡尔哲学,将疼痛视为大脑中“特定”外周疼痛感受器受到刺激的结果。 在 20 世纪,发生了两种对立理论之间的科学斗争,即特异性理论和模式理论。 笛卡儿的“特异性理论”将疼痛视为具有自身器官的感觉输入的一种特定的独立模态,而“模式理论”则认为疼痛是由非特异性受体的强烈刺激引起的。 2 1965 年,Wall 和 Melzack 的 3疼痛门理论为疼痛感知受感觉反馈和中枢神经系统调节的模型提供了证据。 大约在同一时间,疼痛理论的另一个巨大进步是发现了阿片类药物的特定作用模式。4 随后,神经影像学和分子医学的最新进展极大地扩展了我们对疼痛的整体理解。
那么这与肩痛有什么关系呢?肩痛是临床常见问题,并且深入了解身体处理疼痛的方式对于最好地诊断和治疗患者的疼痛至关重要。 我们对疼痛处理知识的进步有望解释病理学与疼痛感知之间的不匹配,它们也可能有助于我们解释为什么某些患者对某些治疗没有反应。
疼痛的基本组成部分
外周感觉感受器:机械感受器和“伤害感受器”
人体肌肉骨骼系统中存在多种类型的外周感觉受体。 5 它们可以根据其功能(如机械感受器、温度感受器或伤害感受器)或形态(游离神经末梢或不同类型的封装受体)进行分类。 5 然后可以根据不同类型的受体进一步细分某些化学标记的存在。 不同功能类别的受体之间存在显着重叠,例如
外周疼痛处理:“伤害感受”
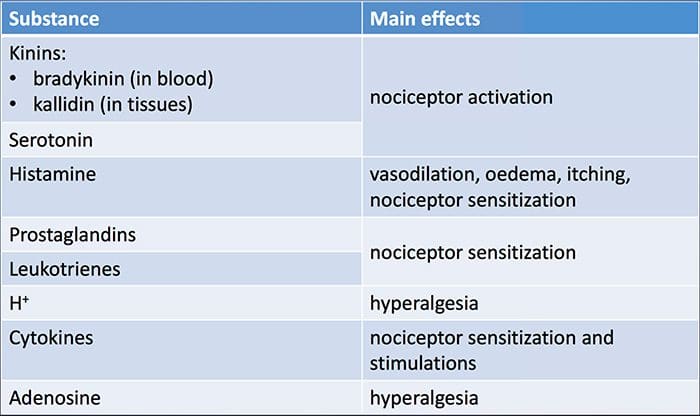
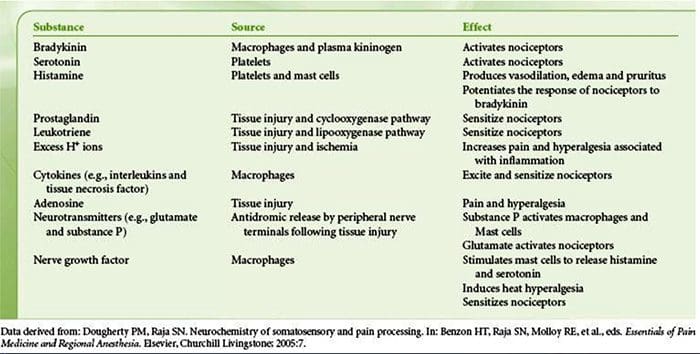
组织损伤涉及受损细胞释放的多种炎症介质,包括缓激肽、组胺、5-羟色胺、ATP、一氧化氮和某些离子(K+ 和 H+)。 花生四烯酸途径的激活导致前列腺素、血栓素和白三烯的产生。 细胞因子,包括白细胞介素和肿瘤坏死因子 α,以及神经营养因子,如神经生长因子 (NGF),也会被释放,并与炎症的促进密切相关。 15 其他物质,如兴奋性氨基酸(谷氨酸)和阿片类药物( endothelin-1) 也与急性炎症反应有关。 16 17 这些药物中的一些可能直接激活伤害感受器,而另一些会引起其他细胞的募集,然后释放更多的促进剂。 18 这种局部过程导致反应性增加伤害性神经元对其正常输入和/或对正常亚阈值输入的响应的募集被称为“外周致敏”。图 1 总结了所涉及的一些关键机制。
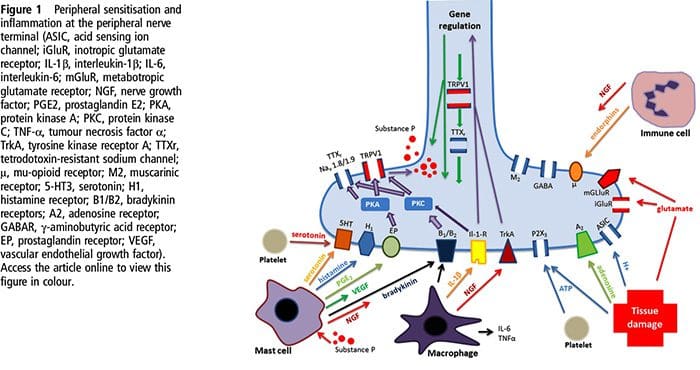 NGF 和瞬时受体电位阳离子通道亚家族 V 成员 1 (TRPV1) 受体在炎症和伤害感受器致敏方面具有共生关系。 发炎组织中产生的细胞因子导致 NGF 产生增加。 19 NGF 刺激肥大细胞释放组胺和血清素 (5-HT3),也使伤害感受器敏感,可能改变 A? 纤维使得更大的比例变得伤害性。 TRPV1 受体存在于初级传入纤维的亚群中,并被辣椒素、热量和质子激活。 TRPV1 受体在传入纤维的细胞体中合成,并被运输到外周和中央终端,在那里它有助于伤害性传入神经的敏感性。 炎症导致 NGF 在外周产生,然后与伤害感受器末端的 1 型酪氨酸激酶受体结合,然后 NGF 被转运到细胞体,导致 TRPV1 转录的上调,从而增加伤害感受器的敏感性。 19 20 NGF 和其他炎症介质也通过多种次级信使途径使 TRPV1 敏感。 许多其他受体,包括胆碱能受体、α-氨基丁酸 (GABA) 受体和生长抑素受体也被认为与外周伤害感受器敏感性有关。
NGF 和瞬时受体电位阳离子通道亚家族 V 成员 1 (TRPV1) 受体在炎症和伤害感受器致敏方面具有共生关系。 发炎组织中产生的细胞因子导致 NGF 产生增加。 19 NGF 刺激肥大细胞释放组胺和血清素 (5-HT3),也使伤害感受器敏感,可能改变 A? 纤维使得更大的比例变得伤害性。 TRPV1 受体存在于初级传入纤维的亚群中,并被辣椒素、热量和质子激活。 TRPV1 受体在传入纤维的细胞体中合成,并被运输到外周和中央终端,在那里它有助于伤害性传入神经的敏感性。 炎症导致 NGF 在外周产生,然后与伤害感受器末端的 1 型酪氨酸激酶受体结合,然后 NGF 被转运到细胞体,导致 TRPV1 转录的上调,从而增加伤害感受器的敏感性。 19 20 NGF 和其他炎症介质也通过多种次级信使途径使 TRPV1 敏感。 许多其他受体,包括胆碱能受体、α-氨基丁酸 (GABA) 受体和生长抑素受体也被认为与外周伤害感受器敏感性有关。
大量炎症介质与肩部疼痛和肩袖疾病特别相关。21-25 虽然一些化学介质直接激活伤害感受器,但大多数会导致感觉神经元本身的变化,而不是直接激活它。 这些变化可能是早期翻译后或延迟转录依赖的。 前者的例子是由膜结合蛋白磷酸化引起的 TRPV1 受体或电压门控离子通道的变化。 后者的例子包括 NGF 诱导的 TRV1 通道产生的增加和钙诱导的细胞内转录因子的激活。
伤害感受的分子机制
疼痛的感觉提醒我们真正的或即将发生的伤害,并触发适当的保护反应。 不幸的是,疼痛往往比它作为警告系统的用处更长,反而变得慢性和衰弱。 这种向慢性期的转变涉及脊髓和大脑内部的变化,但在初级感觉神经元的水平上,疼痛信息的启动也存在显着的调节。 确定这些神经元如何检测产生热、机械或化学性质的疼痛刺激的努力揭示了新的信号机制,并使我们更接近了解促进从急性疼痛转变为持续疼痛的分子事件。
 伤害感受器的神经化学
伤害感受器的神经化学
谷氨酸是所有伤害感受器中主要的兴奋性神经递质。 然而,成人 DRG 的组织化学研究揭示了两大类无髓 C 纤维。
使疼痛恶化的化学传感器
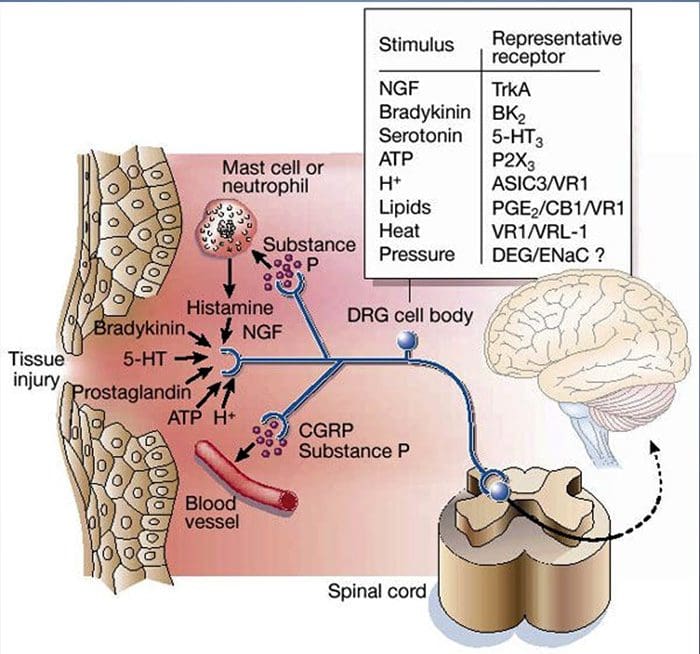
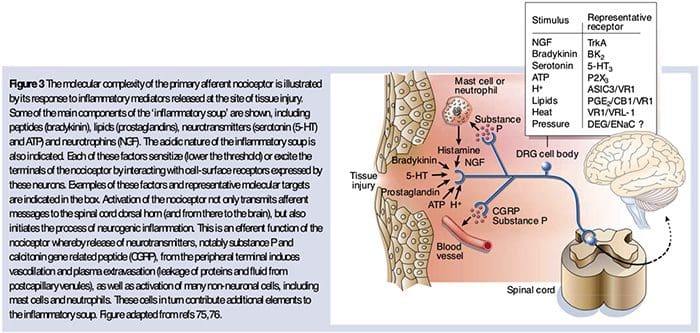
如上所述,伤害通过增加伤害感受器对热和机械刺激的敏感性来增强我们的疼痛体验。 这种现象的部分原因是环境中初级感觉末端和非神经细胞(例如,成纤维细胞、肥大细胞、中性粒细胞和血小板)产生和释放化学介质36(图 3)。 炎症汤的某些成分(例如质子、ATP、11-羟色胺或脂质)可以通过与伤害感受器表面的离子通道相互作用直接改变神经元的兴奋性,而其他成分(例如缓激肽和 NGF)与代谢型受体结合并通过第二信使信号级联介导它们的影响。 在理解这种调节机制的生物化学基础方面已经取得了相当大的进展。
细胞外质子和组织酸中毒
局部组织酸中毒是对损伤的标志性生理反应,相关疼痛或不适的程度与酸化程度密切相关37。 对皮肤施加酸 (pH 5) 会在三分之一或更多支配感受野的多模式伤害感受器中产生持续放电 20。
 疼痛的细胞和分子机制
疼痛的细胞和分子机制
抽象
神经系统检测并解释各种热刺激和机械刺激以及环境和内源性化学刺激物。 当受到强烈刺激时,这些刺激会产生急性疼痛,在持续性损伤的情况下,疼痛传递途径的外周和中枢神经系统组成部分均显示出巨大的可塑性,增强了疼痛信号并产生超敏反应。 当可塑性促进保护性反射时,这可能是有益的,但是当这种变化持续存在时,可能会导致慢性疼痛。 遗传,电生理学和药理学研究正在阐明产生疼痛的有害刺激的检测,编码和调节的分子机制。
简介:急性与持续性疼痛
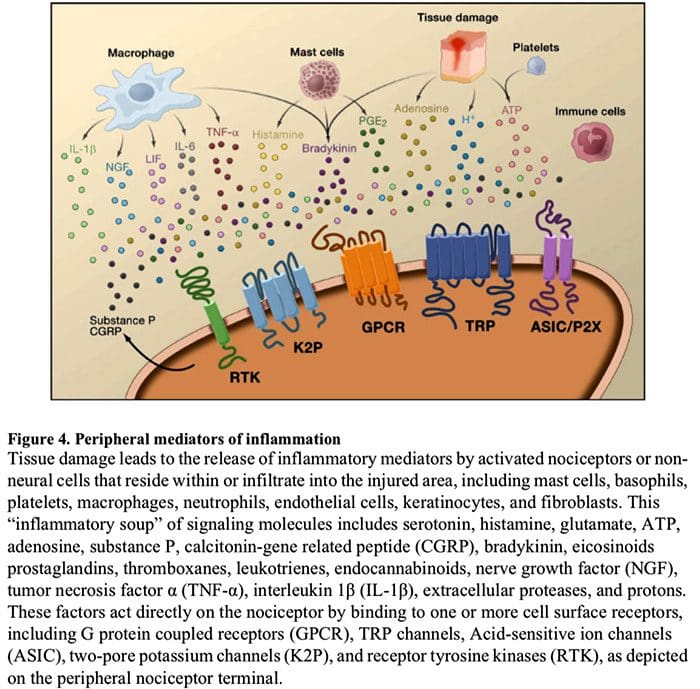
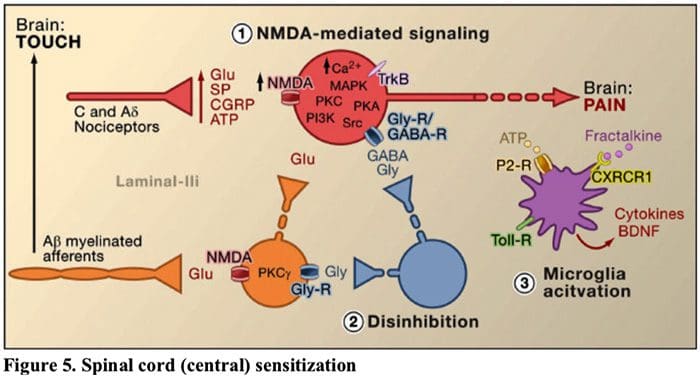 图 5. 脊髓(中央)致敏
图 5. 脊髓(中央)致敏
- 谷氨酸/NMDA 受体介导的致敏作用。�在强烈刺激或持续性损伤后,激活 C 和 A? 伤害感受器释放多种神经递质,包括谷氨酸盐、P 物质、降钙素基因相关肽 (CGRP) 和 ATP,至浅背角(红色)层 I 的输出神经元上。 因此,位于突触后神经元中的通常沉默的 NMDA 谷氨酸受体现在可以发出信号,增加细胞内钙,并激活许多钙依赖性信号通路和第二信使,包括丝裂原活化蛋白激酶 (MAPK)、蛋白激酶 C (PKC) 、蛋白激酶 A (PKA) 和 Src。 这一系列事件将增加输出神经元的兴奋性并促进疼痛信息向大脑的传输。
- 去抑制。�在正常情况下,抑制性中间神经元(蓝色)不断释放 GABA 和/或甘氨酸(Gly)以降低第一层输出神经元的兴奋性并调节疼痛传递(抑制性音调)。 然而,在损伤的情况下,这种抑制可能会丧失,导致痛觉过敏。 此外,去抑制可以使非伤害性有髓 A? 初级传入神经参与疼痛传递电路,因此通常无害的刺激现在被认为是痛苦的。 这部分是通过兴奋性 PKC 的去抑制而发生的? 在内层 II 中表达中间神经元。
- 小胶质细胞激活。�外周神经损伤会促进 ATP 和趋化因子 fractalkine 的释放,从而刺激小胶质细胞。 特别是,小胶质细胞(紫色)上嘌呤能、CX3CR1 和 Toll 样受体的激活导致脑源性神经营养因子 (BDNF) 的释放,其通过激活 lamina I 输出神经元表达的 TrkB 受体,促进增加的兴奋性和对有害和无害刺激(即痛觉过敏和异常性疼痛)的反应增强疼痛。 活化的小胶质细胞还释放出大量细胞因子,如肿瘤坏死因子 ? (TNF?),白细胞介素-1? 和 6 (IL-1?, IL-6),以及其他有助于中枢敏化的因素。
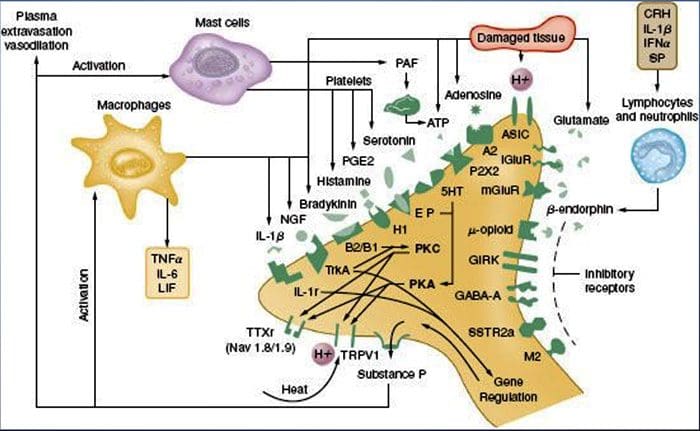
炎症的化学环境
周围神经过敏通常是由神经纤维化学环境中与炎症相关的变化引起的(McMahon等,2008)。 因此,组织损伤通常伴随着活化的伤害感受器或非神经细胞(位于肥大细胞,嗜碱性粒细胞,血小板,巨噬细胞,嗜中性粒细胞,内皮细胞,角质形成细胞和成纤维细胞)。 集体。 这些被称为``炎症汤''的因素代表了广泛的信号分子,包括神经递质,肽类(P物质,CGRP,缓激肽),类花生酸和相关脂质(前列腺素,血栓烷,白三烯,内源性大麻素),神经营养蛋白,细胞因子,趋化因子以及细胞外蛋白酶和质子。 值得注意的是,伤害感受器表达一种或多种细胞表面受体,它们能够识别并响应这些促炎或镇痛药中的每一种(图4)。 这种相互作用增强了神经纤维的兴奋性,从而提高了其对温度或触觉的敏感性。
无疑,减轻炎症性疼痛的最常见方法包括抑制炎症性汤成分的合成或积累。 非甾体类抗炎药,例如阿司匹林或布洛芬,就是最好的例证,它通过抑制参与前列腺素合成的环氧合酶(Cox-1和Cox-2)来减轻炎症性疼痛和痛觉过敏。 第二种方法是阻止炎症药在伤害感受器上的作用。 在这里,我们重点介绍一些实例,这些实例可以为周围致敏性的细胞机制提供新的见解,或者构成治疗炎症性疼痛的新治疗策略的基础。
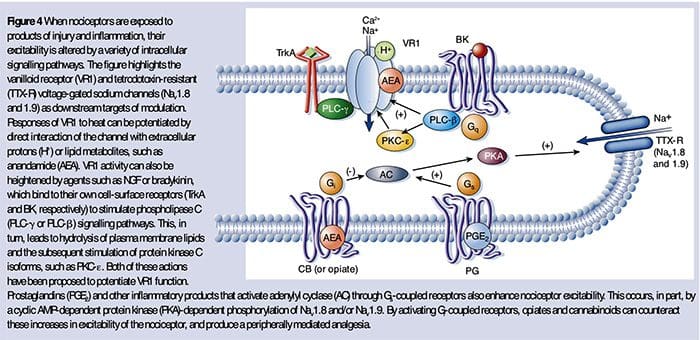
NGF 最为人所知的可能是它作为胚胎发生过程中感觉神经元存活和发育所需的神经营养因子,但在成人中,NGF 也在组织损伤的情况下产生并构成炎症汤的重要成分(Ritner 等人)。等人,2009)。 在其众多细胞靶标中,NGF 直接作用于肽能 C 纤维伤害感受器,后者表达高亲和力 NGF 受体酪氨酸激酶 TrkA 以及低亲和力神经营养因子受体 p75(Chao,2003;Snider 和 McMahon,1998)。 NGF 通过两种时间上不同的机制对热和机械刺激产生深刻的超敏反应。 首先,NGF-TrkA 相互作用激活下游信号通路,包括磷脂酶 C (PLC)、丝裂原活化蛋白激酶 (MAPK) 和磷酸肌醇 3-激酶 (PI3K)。 这导致外周伤害感受器末端靶蛋白的功能增强,最显着的是 TRPV1,导致细胞和行为热敏感性的快速变化(Chuang 等人,2001)。
不管它们的伤害感受机制如何,干扰神经营养因子或细胞因子信号传导已成为控制炎症性疾病或由此产生的疼痛的主要策略。 主要方法包括阻断 NGF 或 TNF-? 用中和抗体作用。 在 TNF-α 的情况下,这在治疗包括类风湿性关节炎在内的多种自身免疫性疾病中非常有效,导致组织破坏和伴随的痛觉过敏的显着减少(Atzeni 等,2005)。 由于 NGF 对成人伤害感受器的主要作用发生在炎症环境中,这种方法的优点是痛觉过敏会降低而不影响 正常的痛觉. 事实上,抗 NGF 抗体目前正处于治疗炎症性疼痛综合征的临床试验中(Hefti 等,2006)。
谷氨酸/ NMDA受体介导的致敏作用
急性疼痛的信号是从伤害感受器的中央末端释放谷氨酸,在二阶背角神经元中产生兴奋性突触后电流 (EPSC)。 这主要通过激活突触后 AMPA 和离子型谷氨酸受体的红藻氨酸亚型而发生。 突触后神经元中亚阈值 EPSC 的总和最终将导致动作电位放电并将疼痛信息传递到更高阶的神经元。
其他研究表明,投射神经元本身的变化有助于去抑制过程。 例如,周围神经损伤会显着下调 K+-Cl- 协同转运蛋白 KCC2,这对于维持跨质膜的正常 K+ 和 Cl- 梯度至关重要(Coull 等,2003)。 下调在 I 层投射神经元中表达的 KCC2 会导致 Cl- 梯度的变化,从而使 GABA-A 受体的激活去极化,而不是使 I 层投射神经元超极化。 反过来,这将增强兴奋性并增加疼痛传递。 事实上,大鼠体内 KCC2 的药理学阻断或 siRNA 介导的下调会诱导机械性异常性疼痛。

分享电子书
来源:
为什么我的肩膀会痛? 肩痛的神经解剖学和生化基础综述
本杰明·约翰·弗洛伊德·迪恩、斯蒂芬·爱德华·格威林、安德鲁·乔纳森·卡尔
疼痛的细胞和分子机制
Allan I. Basbaum1、Diana M. Bautista2、Gre?gory Scherrer1 和 David Julius3
1加州大学解剖学系,旧金山 94158
2加州大学伯克利分校分子与细胞生物学系 CA 94720 3加州大学旧金山分校生理学系 94158
伤害感受的分子机制
大卫朱利叶斯*和艾伦 I. 巴斯鲍姆。
*美国加利福尼亚大学旧金山分校细胞和分子药理学系、解剖学和生理学系和 WM 凯克基金会综合神经科学中心,旧金山,加利福尼亚州 94143,美国(电子邮件: julius@socrates.ucsf.edu)
本文中关于“疼痛的生化” 无意取代与合格的医疗保健专业人员或执业医师的一对一关系,也不是医疗建议。我们鼓励您根据您的研究以及与合格的医疗保健专业人员的合作关系做出医疗保健决策。
我们的信息范围 仅限于脊椎按摩疗法、肌肉骨骼、物理药物、健康、病因 内脏障碍 在临床表现、相关的躯体内脏反射临床动力学、半脱位复合体、敏感的健康问题和/或功能医学文章、主题和讨论中。
我们提供并呈现 临床合作 与来自不同学科的专家。 每位专家均受其专业执业范围和许可管辖范围的约束。 我们使用功能性健康和保健方案来治疗和支持肌肉骨骼系统损伤或疾病的护理。
我们的视频、帖子、主题、主题和见解涵盖与我们的临床实践范围相关并直接或间接支持我们的临床事务、问题和主题。*
我们的办公室已合理尝试提供支持性引文,并已确定支持我们帖子的相关研究或研究。 我们应要求向监管委员会和公众提供支持研究的副本。
我们了解我们承保的事项需要额外解释它如何有助于特定护理计划或治疗方案; 因此,要进一步讨论上述主题,请随时询问 亚历克斯·希门尼斯博士,哥伦比亚特区, 或与我们联系 915-850-0900.
我们在这里为您和您的家人提供帮助。
祝福
Alex Jimenez博士 直流电 麻省理工学院, 注册护士*, 科技委, 国际气候变化联合会*, 国际金融中心*, ATN*
电子邮件: Coach@elpasofunctionalmedicine.com
获得脊椎按摩疗法医生 (DC) 执照 德州 & 新墨西哥*
德克萨斯州特区许可证# TX5807,新墨西哥州 DC 许可证# NM-DC2182
获得注册护士 (RN*) 执照 in 佛罗里达
佛罗里达州执照 注册护士执照 # RN9617241 (控制编号 3558029)
紧凑状态: 多州许可证: 授权执业于 40个州*
Alex Jimenez 博士 DC、MSACP、RN* CIFM*、IFMCP*、ATN*、CCST
我的数字名片





