抑郁症是美国最常见的心理健康问题之一。 当前的研究表明,抑郁症是遗传,生物学,生态和心理方面的综合结果。 抑郁症是世界范围内的一种主要精神疾病,对社会造成重大的经济和心理压力。 幸运的是,即使是最严重的情况,也可以治疗抑郁症。 越早开始治疗,越有效。
然而,结果是,需要鲁棒的生物标志物,其将有助于改善诊断,以加速针对患有该疾病的每个患者的药物和/或药物发现过程。 这些是客观的,外围的生理指标,这些指标的存在可用于预测抑郁症的发作或存在的可能性,根据严重程度或症状进行分层,指示预测和预后或监测对治疗干预措施的反应。 下一篇文章的目的是展示有关发现各种生物的最新见解,当前挑战和未来前景。 生物标志物 以及如何帮助他们改善诊断和治疗。
抑郁症的生物标志物:最新见解,当前挑战和未来前景
抽象
大量的研究已经暗示了数百种抑郁症的生物标志物,但尚未完全阐明其在抑郁症中的作用或确定哪些异常患者以及如何利用生物学信息来增强诊断,治疗和预后。 缺乏进展的部分原因是抑郁症的性质和异质性,再加上研究文献中方法学的异质性以及大量具有潜力的生物标志物,其表达常常根据许多因素而变化。 我们回顾了现有的文献,这些文献表明涉及炎症,神经营养和代谢过程以及神经递质和神经内分泌系统组成的标志物是很有希望的候选物。 这些可以通过遗传和表观遗传学,转录组学和蛋白质组学,代谢组学和神经影像学评估来测量。 现在需要使用新颖的方法和系统的研究程序来确定是否以及哪些生物标记物可用于预测对治疗的反应,将患者对特定治疗进行分层以及制定新干预措施的目标。 我们得出结论,通过进一步发展和扩展这些研究途径来减轻抑郁症的负担有很大希望。
关键词: 情绪障碍,重度抑郁症,炎症,治疗反应,分层,个性化药物
介绍
心理健康和情绪障碍的挑战
尽管精神病学与疾病相关的负担比任何其他医学诊断类别都大,1在包括研究经费2和出版3在内的许多领域,身心健康之间仍然存在明显的自尊差距。3精神卫生面临的困难是缺乏关于分类,诊断和治疗的共识,这是由于对这些疾病的潜在过程的不完全了解所致。 这在情绪障碍中尤为明显,该类别包括精神健康中最大的负担。60最普遍的情绪障碍,重度抑郁症(MDD)是一种复杂的异质性疾病,其中多达4%的患者可能会经历在某种程度上延长或加重发作的治疗抵抗力。5对于情绪障碍,以及在更广泛的精神健康领域,通过在诊断类别内(以及跨诊断类别)发现强而均一的亚型,可能会改善治疗效果。可以分层。 认识到这一点,描述功能亚型的全球计划正在研究中,例如研究领域标准。6人们认为,生物标志物是精神障碍亚型的优先考虑对象。XNUMX
改善对抑郁症治疗的反应
尽管针对重度抑郁症的治疗选择范围广泛,但即使按照共识性指南并采用基于测量的护理接受最佳的抗抑郁药治疗,MDD患者中也只有约三分之一的患者实现了缓解,并且每次新疗法的治疗反应率似乎都下降了.7此外,从长期来看,抗药性抑郁症(TRD)与功能损害,死亡率,发病率以及反复发作或慢性发作有关。8,9因此,在任何临床阶段获得治疗反应的改善将为患者提供更广泛的益处。抑郁症的总体结果。 尽管TRD造成了沉重的负担,但该领域的研究仍然很少。 尽管先前尝试过,TRD的定义仍未标准化:4有些标准仅要求一项治疗试验无法将症状评分降低50%(通过对抑郁症严重程度的有效评估),而其他标准则要求未实现完全缓解或对至少两种经过充分试验的不同类别的抗抑郁药无反应或被认为是TRD。4,10此外,通过在失败的治疗方法中增加严重程度和慢性的关键临床特征,可以提高治疗抵抗性的分期和预测.9,11然而,这种定义上的不一致使得解释TRD的研究文献变得更加复杂。
为了改善对治疗的反应,明确无反应的预测危险因素显然是有帮助的。 已经确定了一些TRD的一般预测指标,包括先前发作后缺乏完全缓解,合并症焦虑,自杀性和抑郁的早期发作以及人格(特别是低外向性,低奖赏依赖性和高度神经质)和遗传因素[12]。综述分别综合了抑郁症的药物治疗和心理治疗的证据,证实了这些发现。 抗抑郁药和认知行为疗法的疗效近似可比[13],但由于其不同的作用机制,可能预期其反应的预测因子也不同。 虽然早年创伤长期以来一直与较差的临床结果和对治疗的反应减少相关,14早期迹象表明,有儿童期创伤史的人对心理的反应可能比药物疗法更好。15尽管如此,不确定性仍然盛行,很少有个性化或治疗分层已达到临床实践16。
这篇综述集中在支持生物标志物作为增强抑郁症治疗反应的潜在有用临床工具的证据上。
生物标志物:系统和来源
生物标志物为识别各种干预反应的预测因素提供了潜在的目标。19迄今为止的证据表明,反映炎症,神经递质,神经营养,神经内分泌和代谢系统活动的标志物可能能够预测目前处于抑郁状态的个体的心理和身体健康状况,但发现之间存在很多不一致之处。20在本综述中,我们重点介绍了这五个生物系统。
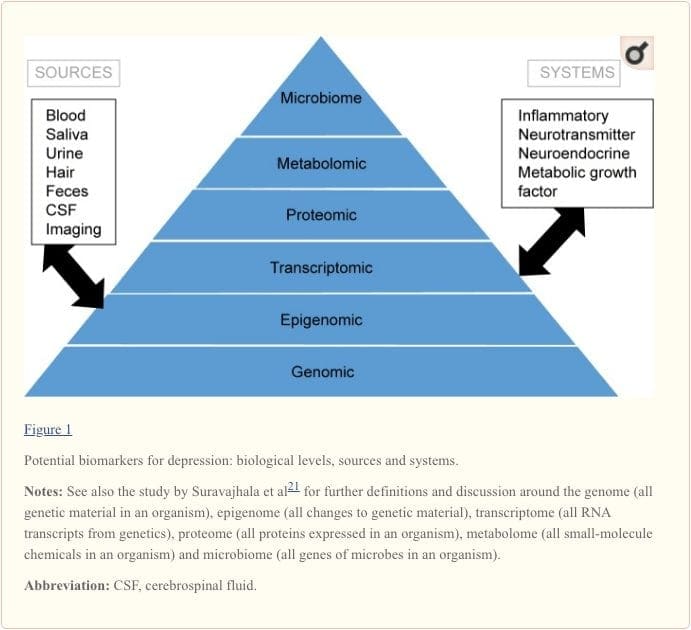
为了全面了解分子途径及其在精神疾病中的作用,现在认为评估多种生物学水平很重要,这被通称为``组学''方法21.图1描绘了不同的方法。可以评估五个系统中每个系统的生物学水平,以及可以进行这些评估的标记物的潜在来源。 但是,请注意,虽然可以在每个组学级别检查每个系统,但是最佳测量源显然在每个级别上都不同。 例如,神经影像学提供了一个间接评估大脑结构或功能的平台,而血液中的蛋白质检查则可以直接评估标记物。 转录组学22和代谢组学23越来越流行,可提供潜在的大量标记物评估,人类微生物组计划正在尝试鉴定人体内所有微生物及其遗传组成。24新型技术正在增强我们对这些微生物的测量能力,包括通过其他途径; 例如,现在可以在头发或指甲(提供慢性适应症)或汗液(提供连续测量)25以及血液,脑脊液,尿液和唾液中测定激素(例如皮质醇)XNUMX。
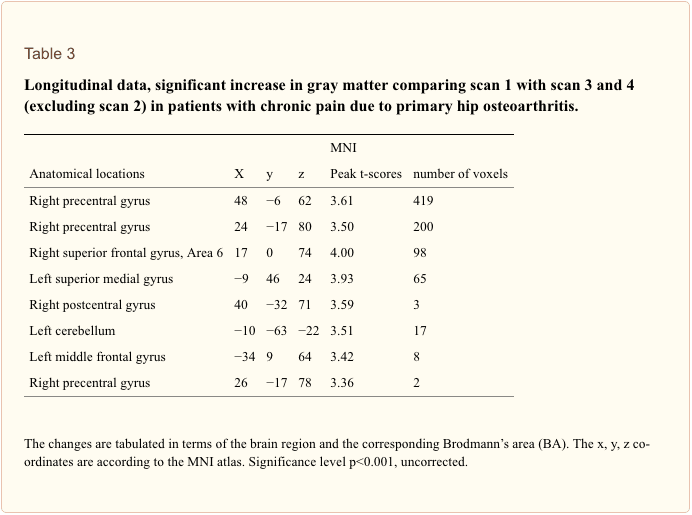
考虑到抑郁症的推定来源,水平和系统的数量,具有翻译潜力的生物标志物的规模如此之大就不足为奇了。 特别是,当考虑标记之间的相互作用时,单独检查单个生物标记可能不会产生有益于改善临床实践的发现。 Schmidt等人[26]提出了使用生物标志物检测小组的建议,随后,Brand等人[27]根据MDD的先前临床和临床前证据概述了一个检测组的草案,确定了16种强生物标志物靶标,每个靶标很少是单一标志物。 它们包括减少的灰质体积(海马,前额叶皮层和基底神经节区域),昼夜节律变化,皮质醇过多和下丘脑-垂体-肾上腺(HPA)轴过度活化的其他表现,甲状腺功能障碍,多巴胺减少,去甲肾上腺素或5-羟吲哚乙酸,谷氨酸增加,超氧化物歧化酶和脂质过氧化增加,环状3',5'-单磷酸腺苷减弱和丝裂原激活的蛋白激酶途径活性,促炎细胞因子增加,色氨酸,犬尿氨酸,胰岛素改变和特定的遗传多态性。 这些标记尚未达成共识,可以通过各种方式进行衡量; 显然,有重点和系统的工作必须解决这一艰巨的任务,以证明其临床益处。
本评论的目的
作为有意的广泛评论,本文旨在确定抑郁症中生物标志物研究的总体需求以及生物标志物在何种程度上具有真正的转化潜力来增强对治疗的反应。 我们首先讨论该领域最重要,最令人兴奋的发现,并引导读者进行与相关标志物和比较有关的更具体的评论。 我们结合证据和减少抑郁症负担的需求,概述了当前面临的挑战。 最后,我们展望了应对当前挑战的重要研究途径及其对临床实践的影响。
最近的见解
在过去半个世纪中,对抑郁症患者寻找临床有用的生物标志物的研究已经进行了广泛的研究。 最常用的治疗方法是从单胺抑郁症理论中得出的。 随后,神经内分泌假说引起了人们的广泛关注。 近年来,最丰富的研究围绕着抑郁症的炎症假说。 但是,大量相关的评论文章都集中在所有五个系统上。 有关跨生物标志物系统的最新见解,请参见表1和下表。 虽然在许多水平上进行了测量,但血液来源的蛋白质已得到最广泛的检查,并提供了一种方便,经济高效且比其他来源更接近翻译潜力的生物标志物来源。 因此,更详细地介绍了血液中循环的生物标记。
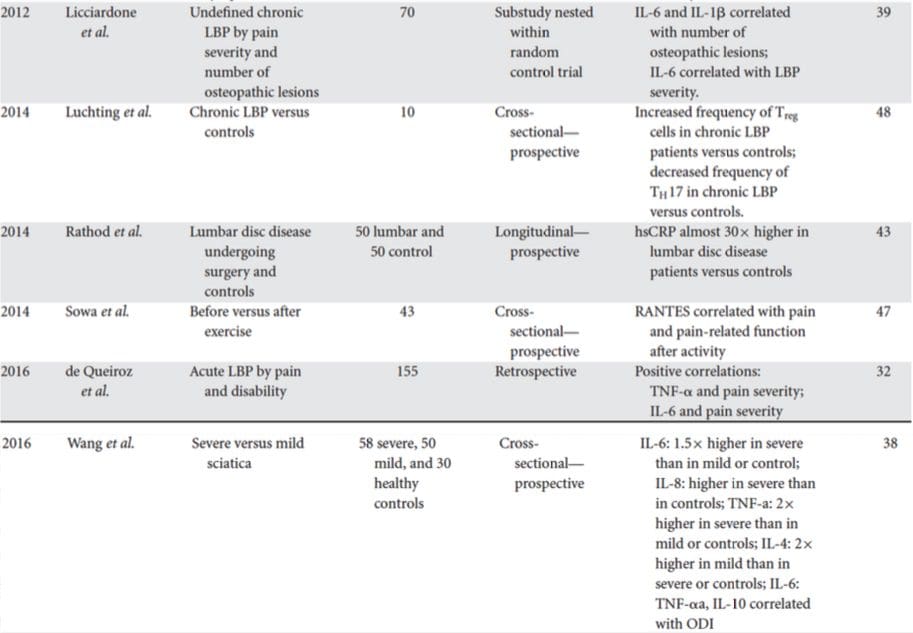
在最近的系统评价中,Jani等[20]检查了与治疗结果相关的抑郁症外周血生物标志物。 在仅纳入的14项研究中(搜索至2013年初),研究了36种生物标志物,其中至少12项研究是心理或身体反应指数的重要预测指标。 被确定为潜在的无反应危险因素的包括炎症蛋白:低白介素(IL)-12p70,淋巴细胞与单核细胞计数之比; 神经内分泌标志物(地塞米松不抑制皮质醇,高循环皮质醇,减少促甲状腺激素); 神经递质标记物(低血清素和去甲肾上腺素); 代谢(低密度脂蛋白胆固醇低)和神经营养因子(减少S100钙结合蛋白B)。 除此之外,其他综述还报道了其他生物标志物与治疗结果之间的关联性19,28-30。随后各节和表2中概述了每个系统中的推定标志物的简要说明。
抑郁症的炎症结果
自从史密斯(Smith)在开创性的论文中概述了巨噬细胞假说以来,[31]这项既有文献发现抑郁症患者各种促炎性标志物的水平有所提高,已得到广泛的评论32。荟萃分析中评估了十二种炎症蛋白,比较了抑郁症和健康控制人群37 38
IL-6(在所有荟萃分析中P <0.001;包括31个研究)和CRP(P <0.001; 20个研究)在抑郁症中频繁且可靠地升高。40在早期研究中发现肿瘤坏死因子α(TNFα)升高。 (P <0.001),38,但是考虑到最近的研究(31项研究)时,实质性异质性并没有定论。40IL-1? 与抑郁症的关系更为不确定,荟萃分析提示抑郁症的水平较高(P = 0.03),41仅在欧洲研究中为高水平42,与对照组无差异。40尽管如此,最近的一篇文章建议对IL- IL-1α升高的极显着作用支持了44?1。 核糖核酸预示着对抗抑郁药的不良反应; 45上述其他发现与循环血源性细胞因子有关。 一项荟萃分析显示,趋化因子单核细胞趋化蛋白-1在抑郁的参与者中升高。39抑郁患者与对照组之间的白介素IL-2,IL-4,IL-8,IL-10和γ干扰素之间无显着差异。荟萃分析的水平,但仍显示出治疗改变的潜力:据报道前瞻性和横断面严重抑郁症患者IL-8水平升高,46治疗期间IL-10和干扰素γ的变化模式不同在早期反应者与无反应者之间发生了47,而IL-4和IL-2的下降与症状缓解一致[48]。在荟萃分析中,IL-6,IL-1?,IL- 10和CRP.43,49,50另外,TNF? 可能仅在应答者中随治疗而减少,并且复合标记指数可能指示随后对治疗无反应的患者炎症增加。43然而,值得注意的是,几乎所有研究炎症蛋白和治疗反应的研究均采用药理学治疗试验。 因此,治疗期间至少一些炎症改变很可能归因于抗抑郁药。 尚未确定各种抗抑郁药的确切炎症作用,但使用CRP水平的证据表明,个体对基于基线炎症的特定治疗反应不同:Harley等[51]报道治疗前CRP升高,预示着对心理疗法(认知行为或人际交往)的反应较差。心理疗法),但对去甲替林或氟西汀的反应良好; Uher等[52]复制了去甲替林的这一发现,并确定了依他普仑的相反作用。 相反,Chang等[53]发现氟西汀或文拉法辛的早期反应者的CRP高于无反应者。 此外,TRD和高CRP的患者对TNF的反应更好。 拮抗剂英夫利昔单抗水平高于正常范围54。
总之,有证据表明,即使控制诸如体重指数(BMI)和年龄之类的因素,大约三分之一的抑郁症患者的炎症反应仍然异常。55,56然而,炎症系统极其复杂,并且有许多生物标志物代表该系统的不同方面。 最近,其他新型细胞因子和趋化因子已产生抑郁症异常的证据。 这些包括:巨噬细胞抑制蛋白1a,IL-1a,IL-7,IL-12p70,IL-13,IL-15,嗜酸性粒细胞趋化因子,粒细胞巨噬细胞集落刺激因子,57 IL-5,58、16,59 IL-17,60、4,61 IL- 62单核细胞趋化蛋白-3胸腺和激活调节趋化因子,63趋化因子-10,64,TNFb,65γ干扰素诱导的蛋白66血清淀粉样蛋白A,1.67可溶性细胞内粘附分子XNUMX和可溶性血管细胞粘附分子XNUMX
抑郁症中生长因子的发现
鉴于非神经营养生长因子(例如与血管生成有关的那些因子)的潜在重要性,我们在生长因子的更广泛定义下指代神经源性生物标志物。
脑源性神经营养因子(BDNF)是其中最常研究的。 多项荟萃分析显示,血清BDNF蛋白的衰减似乎与抗抑郁药治疗同时增加.68这些分析中的最新结果表明,在最严重的抑郁症患者中这些BDNF畸变更为明显,但抗抑郁药似乎可以缓解这些疾病。 71即使在没有临床缓解的情况下,仍可增加这种蛋白质的水平。70BDNF的研究尚未成熟的BDNF进行研究,但两者在功能上(就其对酪氨酸受体激酶B受体的影响而言)似乎有所不同。有证据表明,虽然成熟的BDNF可能会降低抑郁症,但proBDNF可能会过量生产。72在荟萃分析中,据报道外周评估的神经生长因子抑郁症患者的神经生长因子低于对照组,但尽管抗抑郁治疗可能不会改变73在胶质细胞的荟萃分析中也有类似的报道。系源性神经营养因子74
血管内皮生长因子(VEGF)与VEGF家族的其他成员(例如VEGF-C,VEGF-D)一起在促进血管生成和神经发生中起作用,并有望降低抑郁症。75尽管证据不一致,但两项荟萃分析最近表明与对照组相比,抑郁症患者的血液中VEGF升高(横跨16个研究; P <0.001)。76,77然而,TRD78中发现低VEGF,并且较高的水平已预示着对抗抑郁药治疗无反应。79目前尚不清楚为什么VEGF蛋白水平会升高,但可能部分归因于抑郁状态下的促炎活性和/或血脑屏障通透性增加,从而导致脑脊液表达降低80.VEGF与治疗反应之间的关系尚不清楚; 一项最新研究发现,尽管抗抑郁药治疗降低,但血清VEGF或BDNF与反应或抑郁严重程度之间没有关系。81胰岛素样生长因子-1是神经源性功能的另一个因子,在抑郁症中可能增加,反映了抑郁症的失衡。 82,83碱性成纤维细胞生长因子(或FGF-2)是成纤维细胞生长因子家族的成员,在抑郁症患者中似乎比对照组高。84 有人发现,这种蛋白质在MDD中的含量低于健康人,但与抗抑郁药相比,其含量进一步降低了。85
在抑郁症中尚未得到充分探索的其他生长因子包括酪氨酸激酶2和可溶的fms样酪氨酸激酶-1(也称为sVEGFR-1),它们与VEGF协同作用,酪氨酸激酶受体(与BDNF结合)可能会减弱86胎盘生长因子也是VEGF家族的一部分,但据我们所知,尚未在系统性抑郁症样本中进行过研究。
抑郁症的代谢生物标志物发现
与代谢性疾病相关的主要生物标志物包括瘦素,脂联素,生长素释放肽,甘油三酸酯,高密度脂蛋白(HDL),葡萄糖,胰岛素和白蛋白。87许多与抑郁症之间的关联已得到综述:瘦素88和ghrelin89在抑郁症中较低可能会比抗抑郁药的治疗效果好或缓解。 抑郁症患者的胰岛素抵抗可能会增加,尽管少量也可能增加。90包括高密度脂蛋白胆固醇在内的许多抑郁症患者(包括无合并症的患者)的血脂谱似乎发生了改变,尽管这种关系很复杂并且需要进一步阐明。91此外,评论中报道了抑郁症中的高血糖92和低白蛋白血症93。
使用小分子的代谢组学研究整体代谢状态的研究正变得越来越频繁,希望找到一种针对精神疾病的强大生化特征。 在最近的一项使用人工智能建模的研究中,一组代谢物说明葡萄糖脂信号的增加可以高度预测MDD诊断,94支持先前的研究95。
抑郁症中神经递质的发现
尽管人们对抑郁症中单胺的关注已经获得了相对成功的治疗方法,但尚未发现有可靠的神经递质标记物可以根据抗抑郁药单胺靶标的选择性来优化治疗效果。 最近的工作指出5-羟色胺(1-羟色胺)96A受体对于抑郁症的诊断和预后均具有潜在的重要性,尚待开发新的遗传和影像学技术。5有针对性的针对5-羟色胺的新疗法。 例如,使用97-羟基色氨酸的缓释给药。98多巴胺传递的增加与其他神经递质相互作用,从而改善诸如决策和动机之类的认知结果。5同样,神经递质谷氨酸,去甲肾上腺素,组胺和5-羟色胺可能相互作用并激活作为与抑郁相关的压力反应的一部分; 这可能会通过驱油降低99-羟色胺的产量。 最近的综述提出了这一理论,并提出在TRD中,可以通过针对多种神经递质的多模式治疗来逆转(并恢复100-HT)。3有趣的是,血清素的增加并不总是与治疗性抗抑郁药同时发生[4]。 ,通常发现神经递质代谢物,例如去甲肾上腺素的101,102-甲氧基-102,103-羟苯基乙二醇或多巴胺的高香草酸,随着抗抑郁药的治疗而减少抑郁症的同时增加XNUMX或这些代谢物的低水平预示着对SSRI处理XNUMX
神经内分泌在抑郁症中的发现
皮质醇是在抑郁症中最常见的HPA轴生物标志物。 许多评论集中在对HPA活动的各种评估上。 总的来说,这些表明抑郁症与高皮质醇血症有关,并且皮质醇的唤醒反应通常会减弱。104,105这是最近对头发中慢性皮质醇水平进行回顾的证据,支持了抑郁症中皮质醇过高的假设,但在其他疾病(例如其他疾病)中存在过低的假设。 106此外,尤其是皮质醇水平升高可能预示着对心理107和抗抑郁药108治疗的反应较差。 从历史上看,前瞻性治疗反应最有希望的神经内分泌标记物是地塞米松抑制试验,地塞米松给药后皮质醇的不抑制与随后缓解的可能性降低有关。 但是,这种现象尚未被认为对于临床应用而言足够牢固。 在抑郁症中不一致地发现了促肾上腺皮质激素释放激素,促肾上腺皮质激素激素和血管加压素的相关标志物过量,而脱氢表雄酮则被减弱。 皮质醇与脱氢表雄酮的比例可作为TRD中相对稳定的标志物升高,并在缓解后持续存在。109神经内分泌激素功能障碍长期以来与抑郁症相关,甲状腺功能减退也可能在情绪低落中起因果作用。110此外,甲状腺反应可通过成功的抑郁症治疗使之正常化111。
在上述内容中,考虑跨系统的信号传导途径也很重要,例如糖原合酶激酶3,丝裂原活化的蛋白激酶和环状腺苷3′,5′-单磷酸,它们参与突触可塑性112,并被抗抑郁药修饰113。跨生物系统的潜在生物标志物候选物尤其是使用神经成像或遗传学方法进行测量。 为应对抑郁和非抑郁人群之间缺乏稳健而有意义的基因组差异114,新颖的遗传学方法(例如多基因评分115或端粒长度116,117)可能更加有用。 其他越来越流行的生物标记物正在利用不同的来源研究昼夜节律或年代生物学标记物。 书法可以通过加速度计提供客观的睡眠和唤醒活动以及休息的评估,并且书法设备可以越来越多地测量其他因素,例如曝光。 与通常使用的患者主观报告相比,这可能对检测更有用,并且可以提供治疗反应的新预测因子。118哪些生物标志物最适合用于翻译用途是一个具有挑战性的问题,下面将对此进行扩展。
当前的挑战
对于所审查的这五个神经生物学系统中的每一个,证据遵循相似的叙述:存在许多在某些方面与抑郁症相关的生物标志物。 这些标记经常以复杂,难以建模的方式相互关联。 证据不一致,很可能有些是其他因素的现象,而有些仅在一部分患者中很重要。 生物标志物可能会通过多种途径发挥作用(例如,那些可预测对治疗的后续反应的标志,那些指示更可能有效的特定治疗的标志或那些因干预而改变的标志,而与临床改善无关)。 需要新的方法来最大化精神病学人群生物学评估的一致性和临床适用性。
生物标志物变异
生物标志物随时间和情况的变化更多地涉及某些类型(例如蛋白质组学)而不是其他类型(基因组学)。 许多人的标准化规范不存在或尚未被广泛接受。 实际上,环境因素对标志物的影响通常取决于遗传成分和人与人之间无法完全解释的其他生理差异。 这使得对生物标志物活性的评估和鉴定生物学异常变得难以解释。 由于潜在的生物标记物的数量,许多尚未与其他相关标记物一起被广泛或完整地测量。
据报道,许多因素会改变情感障碍患者整个生物学系统的蛋白质水平。 以及与研究相关的因素,例如储存时间和条件(可能导致某些化合物降解),这些因素包括一天中的测量时间,种族,运动,119饮食(例如微生物组活动,尤其是在大多数血液生物标志物研究可以做到的情况下) 120吸烟和使用毒品121以及健康因素(例如合并性炎症,心血管疾病或其他身体疾病)。 例如,尽管在抑郁症患者中观察到炎症加剧,但与非抑郁症患者相比,其他方面健康的患者,与免疫共病相关的抑郁症患者与非抑郁症或疾病患者相比,其细胞因子水平往往更高。122一些明显的因素与下面概述了可能与生物标志物,抑郁症和治疗反应之间的关系。
应力。 内分泌反应和免疫反应在应对压力(生理或心理)方面均具有众所周知的作用,尽管在研究中,个体之间这一因素的可变性可能会因当前的情况而加剧,但在生物样本采集时的短暂压力很少在研究中进行测量。抑郁症状。 急性和慢性心理应激源都可作为一种免疫挑战,短期和长期内都会加剧炎症反应。123,124这一发现扩展到了早期生命应激的经历,该经历与成人炎症升高相关,而成人炎症升高与经历的应激无关。 125,126在儿童期的创伤经历中,也仅在目前处于抑郁状态的儿童中报告了炎症加剧。127相反,与抑郁症和抑郁症患者相比,患有抑郁症和儿童期创伤史的人皮质醇对压力的反应可能减弱。无早期生命创伤。128压力诱导的HPA轴改变似乎与认知功能相关,129以及HPA相关基因的抑郁亚型或变异。130压力也对神经发生131有短期和长期的影响,其他神经132目前尚不清楚儿童创伤如何影响抑郁成人的生物标志物s,但生命早期的压力可能会使某些人在成年期遭受持久的压力反应,这种压力反应在心理和/或生物学上都会加剧。
认知功能。 情感障碍患者甚至在未经药物治疗的MDD中,神经认知功能障碍也经常发生。133认知缺陷与治疗抵抗力一起出现累积现象。134从生物学上讲,HPA轴129和神经营养系统135可能在这种关系中起关键作用。 神经递质去甲肾上腺素和多巴胺可能对学习和记忆等认知过程很重要。136炎症反应升高与认知能力下降有关,并可能通过多种机制影响抑郁症发作137和缓解的认知功能。138实际上, Krogh等[139]提出,CRP与认知表现的关系比与抑郁症的核心症状的关系更紧密。
年龄,性别和BMI。 迄今为止,男女之间生物学差异的存在与否以及变化的方向在证据方面尤其有差异。 男女之间神经内分泌激素的变化与抑郁症易感性相互作用。140一项炎症研究的回顾报告指出,控制年龄和性别不会影响患者控制炎症细胞因子的差异(尽管随着年龄的增长,IL-6和抑郁症之间的联系会减少, 41,141在评估年轻样本的研究中,患者和对照组之间的VEGF差异更大,而性别,BMI和临床因素在荟萃分析水平上没有影响这些比较。[77]在先前的炎症和抑郁检查中缺乏对BMI的调整似乎混淆了这些组之间报道的高度显着差异。41明确证明,肿大的脂肪组织可刺激细胞因子的产生,并与代谢标志物密切相关。142因为精神药物可能与魏相关体重增加和更高的BMI,这些都与抑郁症的治疗抵抗力有关,这是需要检查的重要领域。
服药。 许多针对抑郁症的生物标志物研究(横断面和纵向)都收集了非药物参与者的基线标本,以减少异质性。 然而,这些评估中的许多评估是在药物清除期之后进行的,这留下了生理学残留变化的潜在显着混杂因素,而现有的可能对炎症产生不同影响的广泛治疗加剧了这种评估。 一些研究排除了精神药物,但没有其他药物的使用:特别是,口服避孕药在研究参与者中经常被允许使用,而在分析中却没有得到控制,最近已表明会增加激素和细胞因子的水平。143,144一些研究表明抗抑郁药药物会影响炎症反应,34,43,49,145、147、108、148·149 HPA轴,2个神经递质,1和神经营养150活性。 然而,许多潜在的抑郁症治疗方法具有独特而复杂的药理特性,这表明在当前数据的支持下,不同治疗方法可能会产生离散的生物学效应。 从理论上讲,除单胺作用外,靶向血清素的特定药物(即SSRIs)也可能针对炎症反应中的ThXNUMX转变,去甲肾上腺素能抗抑郁药(例如SNRIs)也可能导致ThXNUMX转变。XNUMX目前尚不可能确定单独或联合用药对生物标志物的影响。 这些可能是由其他因素介导的,包括治疗时间长短(很少有试验评估长期用药情况),样品异质性以及未根据治疗反应对参与者进行分层。
异质性
方法论上。 如上所述,研究对象之间以及研究对象之前和之前采取的治疗(和组合)方面的差异(研究之间和内部之间的差异)必将在研究结果中引入异质性,特别是在生物标记研究中。 除此之外,许多其他设计和样本特征也会因研究而异,从而增加了解释和归因结果的难度。 这些包括生物标志物测量参数(例如化验试剂盒)以及在抑郁症中收集,存储,处理和分析标志物的方法。 Hiles等[141]检查了一些有关炎症的文献不一致的来源,发现抑郁症诊断,BMI和合并症的准确性是评估抑郁症和非抑郁症患者外周炎症的最重要因素。
临床。 抑郁人群的广泛异质性已有充分文献记载151,并且是导致研究文献中发现差异的关键因素。 即使在诊断中,异常的生物学特征也可能仅限于随着时间的推移可能不稳定的个体子集。 可以通过心理和生物学因素的组合来识别抑郁症患者的凝聚力亚组。 下面,我们概述了探索亚组以应对生物标志物变异性和异质性构成的挑战的潜力。
抑郁症内的亚型
迄今为止,尚无抑郁症发作或障碍中的同质亚组能够根据症状表现或治疗反应性可靠地区分患者。152存在生物畸变更为明显的亚组将有助于解释先前研究与研究之间的异质性。可以催化实现分层治疗的道路。 Kunugi等[153]根据不同的神经生物学系统在抑郁症中表现出临床相关亚型的作用,提出了四种潜在亚型的集合:患有高皮质醇血症伴忧郁症抑郁症或表现出非典型亚型的皮质醇缺乏症的患者,多巴胺相关的患者亚组表现为快感不足(并可能对阿立哌唑有很好的反应)和以炎症加剧为特征的炎症亚型。 许多关于炎症的文章都指出在抑郁症中存在``炎性亚型''的情况.55,56,154,155炎症升高的临床相关性尚未确定,并且几乎没有直接尝试发现哪些参与者可能构成这一队列。 有人提出非典型抑郁症患者的炎症水平可能高于忧郁症亚型156,这可能与忧郁症和非典型抑郁症亚型的HPA轴发现不符。 TRD37或具有明显躯体症状的抑郁症157也被认为是潜在的炎症亚型,但神经营养(睡眠,食欲,性欲减退),情绪(包括情绪低落,自杀和易怒)和认知症状(包括情感偏见和内)158全部看起来与生物特征有关。 炎性亚型的其他潜在候选人还包括类似疾病行为的症状[159,160]或代谢综合征[158]。
对(低)躁狂症的倾向可能会在生物学上区分患有抑郁症的患者。 现在的证据表明,躁郁症是情绪障碍的一个多方面的群体,发现躁郁症的症状较以前认识的更为广泛。161最近,躁郁症的不准确和/或延迟发现是临床精神病学中的一个主要问题。纠正诊断的平均时间通常超过十年,162并且这种延迟导致更大的严重性和更大的整体疾病费用。163大多数双相情感障碍患者最初表现为一个或多个抑郁发作,单相抑郁是最常见的误诊,可能区分单相和双相抑郁的因素具有重大意义。164在先前的某些MDD生物标志物研究中可能未发现双相谱障碍,并且有证据表明双相和单相depr在HPA轴活性109或炎症方面存在差异165,166。 ession。 但是,这些比较很少,样本量较小,发现的趋势影响不明显,或者招募的人群没有通过诊断很好地表征。 这些研究也没有检查在这些关系中治疗反应性的作用。
双相情感障碍167和治疗抗药性168都不是二分法构建的,而是连续存在的,这增加了亚型鉴定的挑战。 除了分型以外,值得注意的是,在患有其他诊断的患者中类似地发现了抑郁症中观察到的许多生物学异常。 因此,转诊检查也可能很重要。
生物标志物测量挑战
生物标志物的选择。 大量潜在有用的生物标志物在确定哪些标志物以何种方式和为谁牵连方面给心理生物学带来了挑战。 为了增加挑战,对这些生物标志物中的相对较少的人进行了抑郁症的充分研究,并且对于大多数生物标志物在健康和临床人群中的确切作用尚不甚了解。 尽管如此,已经进行了许多尝试来提出有前途的生物标志物组。 除了布兰德(Brand)等人的16种具有潜在潜力的标记物外,27罗普雷斯蒂(Lopresti)等人还概述了另外一组广泛的氧化应激标志物,具有改善治疗反应的潜力.28帕帕科斯塔斯(Papakostas)等人事先定义了一套九种血清标记物验证和复制带有MDD的样品的生物系统(BDNF,皮质醇,II型可溶性TNFα受体,α1抗胰蛋白酶,载脂蛋白CIII,表皮生长因子,髓过氧化物酶,催乳素和抵抗素)。 一旦结合起来,对这些水平的综合测量就能够以80%±90%的准确度区分MDD组和对照组。169我们建议,即使是这些,也不能覆盖该领域的所有潜在候选人。 参见表2对具有抑郁可能性的生物标志物的详尽描述,包括具有证据基础和有前途的新型标志物的生物标志物。
技术。 由于技术的进步,与以前相比,现在有可能(确实,方便)以更低的成本和更高的灵敏度同时测量大量生物标志物。 目前,这种测量多种化合物的能力领先于我们有效分析和解释数据的能力170,随着生物标记物阵列和新标记物(例如代谢组学)的兴起,这种能力将继续发展。 这主要是由于对标记物的确切作用以及它们之间的相互关系缺乏了解,并且对相关标记物如何在个体内部和个体之间的不同生物学水平(例如遗传,转录,蛋白质)之间关联的认识不足。 使用新的分析方法和标准的大数据将有助于解决这一问题,并且正在提出新的方法。 一个例子是开发一种基于通量分析的统计方法,该方法基于网络之间的反应来发现新的潜在代谢标记,并将基因表达与代谢物数据整合起来。171机器学习技术已被应用,并将协助使用生物标记的模型数据来预测具有大数据的研究中的治疗结果。172
聚集生物标志物。 同时检查一系列生物标志物是检查孤立的标志物的一种替代方法,可以为更复杂的生物系统网络或网络提供更准确的观点。26此外,还可以帮助弄清迄今为止文献中的对比证据(尤其是在生物标志物网络中)并充分了解了相互作用),然后可以对生物标记数据进行汇总或建立索引。 一个挑战是确定最佳的执行方法,这可能需要技术和/或新颖的分析技术的增强(请参阅``大数据''部分)。 从历史上看,两个不同的生物标志物之间的比率产生了有趣的发现。109,173很少有人尝试在更大范围内汇总生物标志物数据,例如使用促炎性细胞因子网络的主成分分析进行的那些研究。174在荟萃分析中,促炎性细胞因子已经被广泛使用。转换为每项研究的单项疗效评分,总体显示抗抑郁药治疗前的炎症明显更高,这预示了门诊研究随后的无反应。 复合生物标志物组既是未来研究的挑战,也是寻找可用于改善治疗结果的可靠发现的挑战和机遇。43Papakostas等人的一项研究采用了另一种方法,选择了一组异质性血清生物标志物(炎性, HPA轴和代谢系统)在先前的研究中已被证明在抑郁个体和对照个体之间存在差异,并将其综合为两个独立样本和一个对照组的风险评分有所不同,敏感性和特异性均高于80%169。
大数据。 大数据的使用可能是解决当前围绕异质性,生物标志物变异性,确定最佳标志物并将该领域用于抑郁的转化性应用研究的挑战所必需的。 但是,如上所述,这带来了技术和科学挑战。175健康科学直到最近才开始使用大数据分析,比商业领域晚了十年左右。 但是,随着对精神病学生物学机制的了解,诸如iSPOT-D152和诸如精神病学遗传学联盟176之类的联合体的研究正在不断发展。 在极少的研究中,机器学习算法已开始应用于抑郁症的生物标记物:最近的一项调查收集了来自5,000个生物标记物的250多名参与者的数据; 多次插补数据后,进行了机器学习增强的回归分析,表明有21种潜在的生物标志物。 经过进一步的回归分析,选择了三种生物标志物,它们与抑郁症状(高度可变的红细胞大小,血清葡萄糖和胆红素水平)相关性最强。 作者得出的结论是,大数据可以有效地用于产生假设。177目前正在进行较大的生物标志物表型研究项目,这将有助于推动我们进入抑郁症神经生物学的未来。
前景
生物标志物鉴定
迄今为止,文献中的发现需要在大规模研究中进行复制。 对于新的生物标记物尤其如此,例如,趋化因子胸腺和激活调节的趋化因子以及生长因子酪氨酸激酶2,据我们所知,尚未在临床抑郁和健康对照样品中进行过研究。 大数据研究必须分析全面的生物标志物,并使用复杂的分析技术来充分确定标志物与那些在临床和非临床人群中修饰标志物的因素之间的关系。 此外,主成分分析的大规模复制可能会建立高度相关的生物标记物组,也可能会为生物学精神病学中``复合物''的使用提供信息,这可能会增强未来研究结果的同质性。
均质亚型的发现
关于生物标志物的选择,研究可能暗示的不同潜在途径可能需要多个小组。 综上所述,目前的证据表明,在目前患有抑郁症的亚人群中,生物标志物的分布确实可靠,但发生了深远的变化。 这可以在诊断类别内或在诊断类别之间建立,这将解释一些可以在本文献中观察到的发现不一致之处。 抑郁症中生物标志物网络面板的大型聚类分析可以最有效地帮助量化一个或多个生物亚组。 这将说明人口内部的变异性; 潜在类别分析可能会基于例如炎症表现出不同的临床特征。
对炎症和反应的特殊治疗作用
应该对所有常用的抑郁症治疗药物的具体生物学效应进行全面评估,同时还要考虑到治疗试验的有效性。 这可以使与生物标志物和症状表现有关的构建体以更加个性化的方式预测各种抗抑郁治疗的结果,并且在单相和双相抑郁的背景下都是可能的。 这对于新的潜在治疗方法以及当前指示的治疗方法可能很有用。
前瞻性确定治疗反应
使用以上技术可能会提高预期的抗药性预测能力。 更真实和持久(例如,长期)的治疗反应措施可能对此有所帮助。 评估患者福祉的其他有效指标(例如生活质量和日常功能)可以提供更全面的治疗结果评估,从而可以与生物标志物更紧密地关联。 虽然仅靠生物学活动可能无法将治疗反应者与无反应者区分开,但可以将同时具有社会心理或人口统计学变量的生物标志物测量与生物标志物信息结合起来,以建立治疗反应不足的预测模型。 如果开发了一个可靠的模型来预测反应(针对低迷人群或亚群)并进行回顾性验证,则翻译设计可以在大型对照试验中确定其适用性。
走向分层治疗
目前,抑郁症患者没有系统地接受优化的干预计划。 如果得到验证,则可以采用分层试验设计来测试模型,以预测无反应和/或确定需要在分级护理模型中对患者进行分类的位置。 在不同类型的干预措施中,这在标准化和自然主义治疗环境中都可能有用。 最终,可以开发出一种临床上可行的模型,为个体提供最合适的治疗方法,以识别可能发展为难治性抑郁症的人,并为这些患者提供更好的护理和监测。 被确定为有治疗抗性风险的患者可以接受伴随的心理和药物治疗或联合药物治疗。 作为一个推测性的例子,没有炎症性细胞因子升高的参与者可能被指示接受心理治疗而不是药物治疗,而炎症程度特别高的一部分患者可以接受抗炎药以增强标准治疗。 类似于分层,将来可能会出现个性化的治疗选择策略。 例如,一个特定的抑郁个体可能具有明显高的TNF? 水平,但没有其他生物学异常,并且可以从TNF的短期治疗中受益吗? 54个性化治疗还可能需要在治疗过程中监测生物标志物的表达,以告知可能的干预措施变化,所需的持续治疗时间或检测复发的早期标志物。
新型治疗目标
有大量可能有效治疗抑郁症的潜在疗法,尚未得到充分研究,包括来自其他医学学科的新颖或针对性的干预措施。 一些最受欢迎的靶标是抗炎药,例如塞来昔布(和其他环氧合酶2抑制剂),TNF? 拮抗剂依那西普和英夫利昔单抗,米诺环素或阿司匹林。 这些似乎很有希望。178已研究了抗酮糖皮质激素化合物(包括酮康唑179和甲吡酮180)用于抑郁症,但它们的副作用均存在缺点,甲吡酮的临床潜力尚不确定。 米非司酮181和皮质类固醇氟氢可的松和螺内酯182和地塞米松和氢化可的松183可能在短期内也可有效治疗抑郁症。 靶向谷氨酸N-甲基-d-天冬氨酸受体拮抗剂(包括氯胺酮)可能代表抑郁症的有效治疗。184Omega-3多不饱和脂肪酸会影响炎症和代谢活性,并显示出对抑郁症的某些功效。185他汀类药物可能通过相关的神经生物学途径具有抗抑郁作用186。
这样,抗抑郁药的生化作用(见``药物治疗''一节)已被用于其他学科的临床益处:尤其是胃肠病,神经系统疾病和非特异性症状疾病.188抗抑郁药的抗炎作用可能代表了抗抑郁药的部分机制。这些好处。 还已经提出锂可以通过糖原合酶激酶3途径来减轻炎症反应。189对这些作用的关注可以证明抑郁症生物标志物的信息丰富,而生物标志物又可以代表新药开发的替代标志物。
Alex Jimenez博士的见解
抑郁症是一种精神健康疾病,其特征是严重的症状会影响情绪,包括对活动的兴趣下降。 然而,最近的研究发现,不仅可以使用患者的行为症状来诊断抑郁症,还可以。 研究人员认为,识别易于获得的生物标记物以更准确地诊断抑郁症是改善患者整体健康状况的基础。 举例来说,临床发现表明,患有重性抑郁症或MDD的患者血液中的乙酰L-肉碱或LAC分子水平低于健康对照者。 最终,建立抑郁症的生物标记物可能有助于更好地确定谁有患疾病的风险,并帮助医疗保健专业人员确定抑郁症患者的最佳治疗选择。
结论
文献表明,大约三分之二的抑郁症患者未实现初始治疗的缓解,并且无反应的可能性随着试验治疗次数的增加而增加。 提供无效的疗法会对个人和社会成本产生重大影响,包括持续困扰和福祉,自杀风险,生产力损失和医疗资源浪费。 抑郁症方面的大量文献表明,大量生物标志物具有改善抑郁症患者治疗的潜力。 除了已被广泛研究了数十年的神经递质和神经内分泌标记物外,最近的见解还强调了炎症反应(更广泛地说是免疫系统),代谢和生长因子与抑郁症有重要关系。 但是,过多的对比证据表明,在应用生物标志物研究以改善对抑郁症患者的管理和护理之前,有许多挑战需要解决。 由于生物系统极其复杂,因此同时检查大样本中的多种标记物对于发现个体之间的生物学状态和心理状态之间的相互作用具有相当大的益处。 优化神经生物学参数的测量和抑郁症的临床测量可能有助于增进了解。 这篇综述还强调了检查潜在的改变因素(如疾病,年龄,认知和药物治疗)在收集对抑郁症生物学和治疗抗性机制的连贯理解中的重要性。 某些标志物可能最有可能预测亚组患者的治疗反应或对特定治疗的耐药性,同时对生物学和心理数据进行测量可能会增强前瞻性地识别有不良治疗结果风险的人的能力。 建立生物标志物检测小组对于提高诊断的准确性和预后,以及在抑郁症的最早可行阶段中个体化治疗以及制定有效的新治疗靶点都具有重要意义。 这些暗示可能仅限于抑郁症患者的亚组。 通往这些可能性的途径补充了最近的研究策略,以将临床综合症与潜在的神经生物学底物更紧密地联系起来。6除了减少异质性之外,这还可以促进在身体和精神健康方面朝着自尊的方向发展。 显然,尽管需要大量工作,但建立相关生物标志物与抑郁症之间的关系对于减轻个人和社会的抑郁症负担具有实质性意义。
致谢
本报告代表由位于伦敦南部的美国国立卫生研究院(NIHR)生物医学研究中心以及Maudsley NHS基金会信托基金和伦敦国王学院资助的独立研究。 所表达的观点是作者的观点,不一定是NHS,NIHR或卫生部的观点。
脚注
披露。 AHY在过去3年中因其Astra Zeneca(AZ),Lundbeck,Eli Lilly和Sunovion的演讲而获得酬金。 聘请扬纳(Livanova)的艾尔根(Allergan)和扬森(Janssen)的Sunovion伦贝克(Lundbeck)提供咨询服务; 以及来自Janssen和英国资助机构(NIHR,MRC,Wellcome Trust)的研究资助支持。 在过去三年中,AJC曾获得来自Astra Zeneca(AZ)的演讲酬金,来自Allergan,Livanova和Lundbeck的咨询酬金以及来自Lundbeck和英国资助机构(NIHR,MRC,Wellcome Trust)的研究补助金。
作者报告在这项工作中没有其他利益冲突。
总之, ``尽管许多研究发现了数百种抑郁症生物标志物,但很少有人在抑郁症疾病中确立其作用,或者如何准确利用生物学信息来增强诊断,治疗和预后。 但是,以上文章回顾了其他过程中涉及的生物标志物的可用文献,并将临床发现与抑郁症进行了比较。 此外,有关抑郁症生物标志物的新发现可能有助于更好地诊断抑郁症,以便进行更好的治疗。 国家生物技术信息中心(NCBI)引用的信息。 要讨论主题,请随时询问Jimenez博士或通过以下方式与我们联系。915-850-0900 。
由Alex Jimenez博士策划
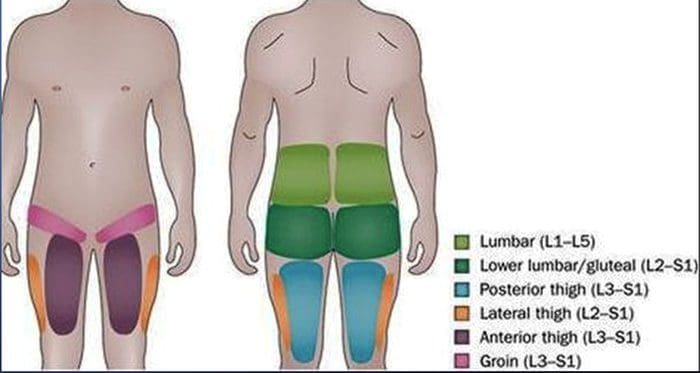
附加主题:腰痛
背疼 是造成残障和世界各地缺勤时间最普遍的原因之一。 事实上,背痛被认为是第二次就诊最常见的原因,仅次于上呼吸道感染。 大约80%的人口在一生中至少会经历某种背痛。 脊柱是由骨骼,关节,韧带和肌肉以及其他软组织组成的复杂结构。 因此,受伤和/或病情加重,例如 椎间盘突出 ,最终会导致背部疼痛的症状。 运动伤害或汽车事故伤害通常是造成背痛的最常见原因,但是,有时最简单的动作可能会产生痛苦的结果。 幸运的是,诸如脊椎治疗等替代疗法可以通过使用脊柱调节和手动操作来帮助缓解背部疼痛,最终改善疼痛缓解。
VIDEO
VIDEO












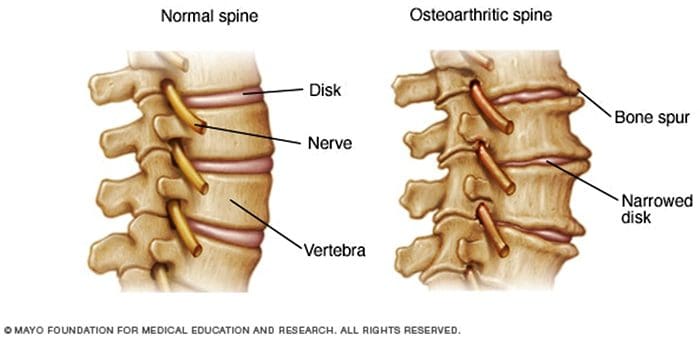
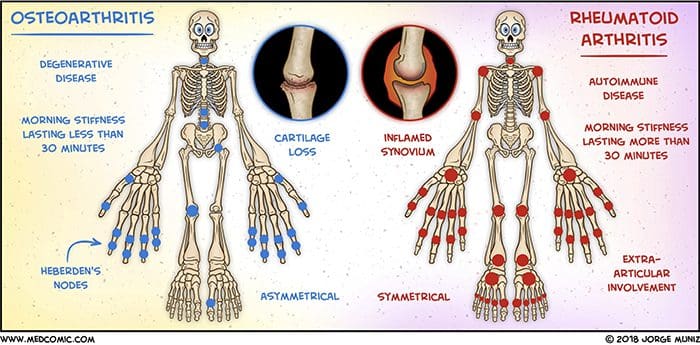 抽象
抽象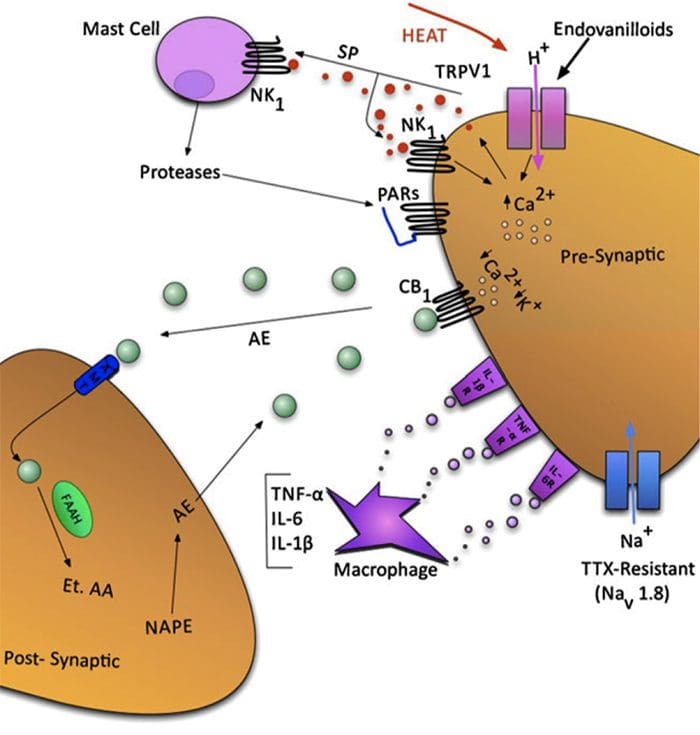








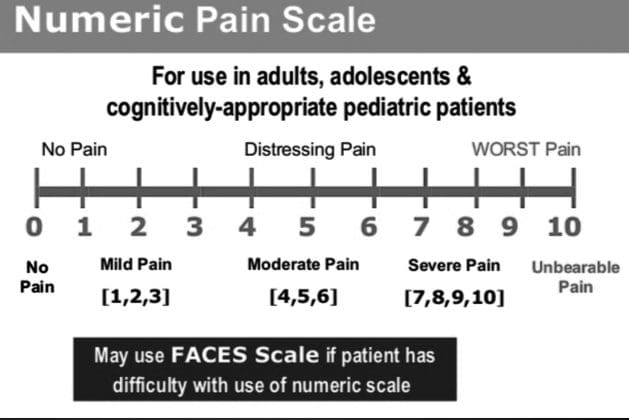
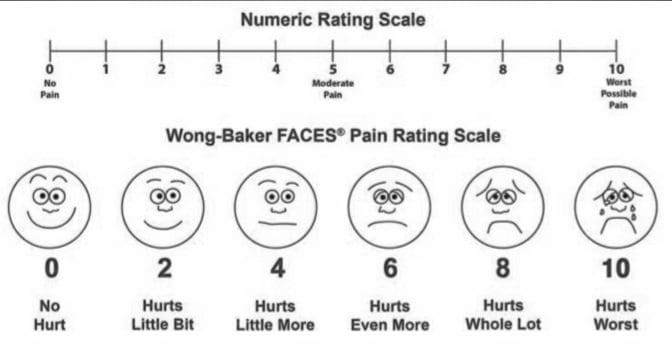










 参考文献:
参考文献:

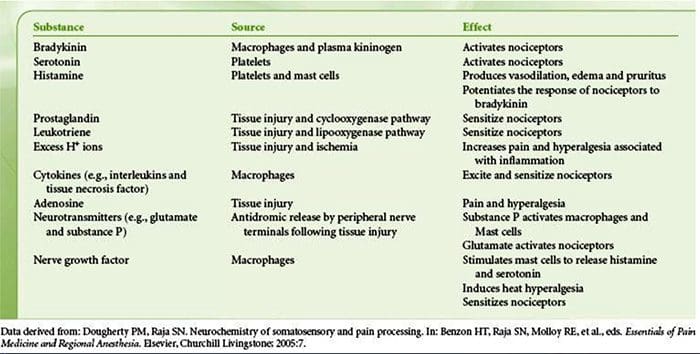
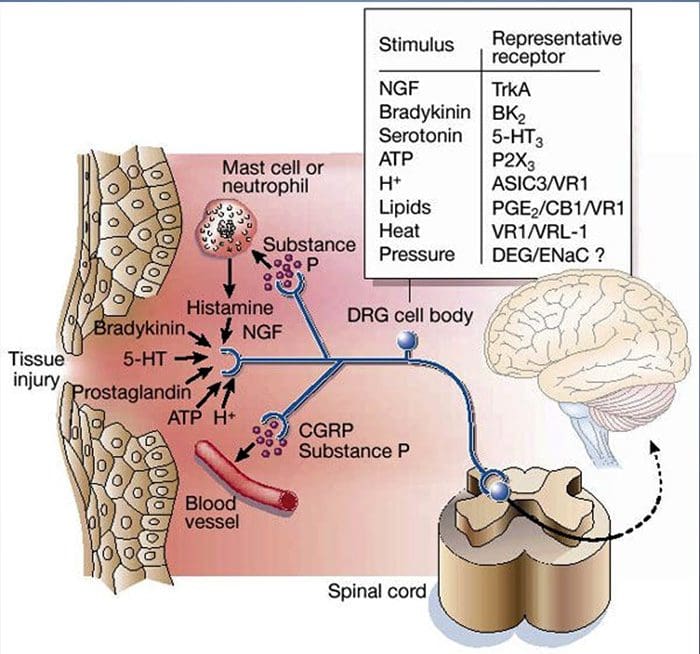
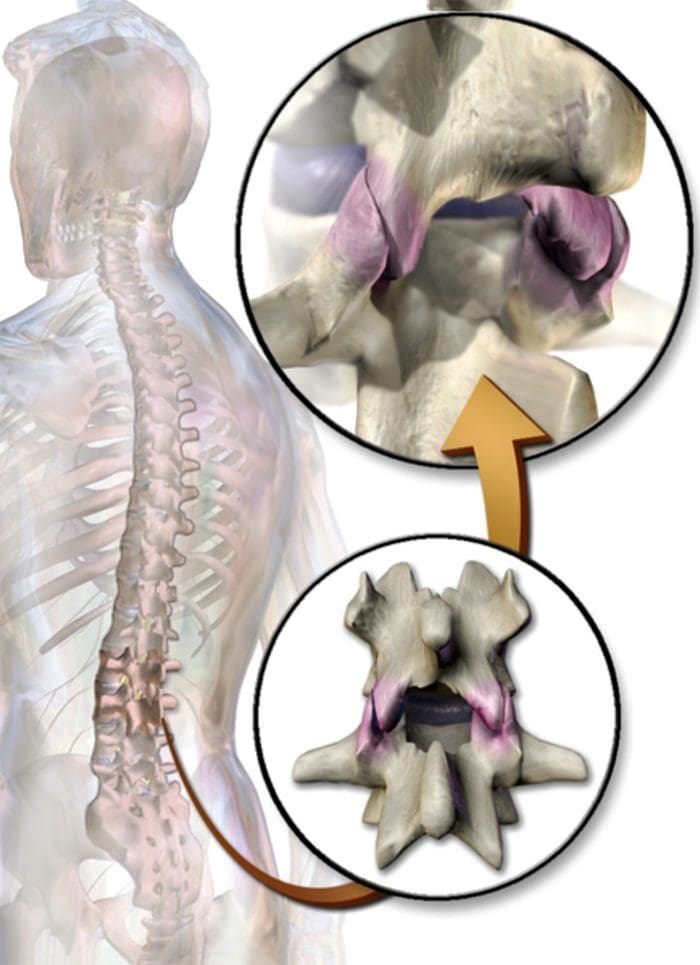
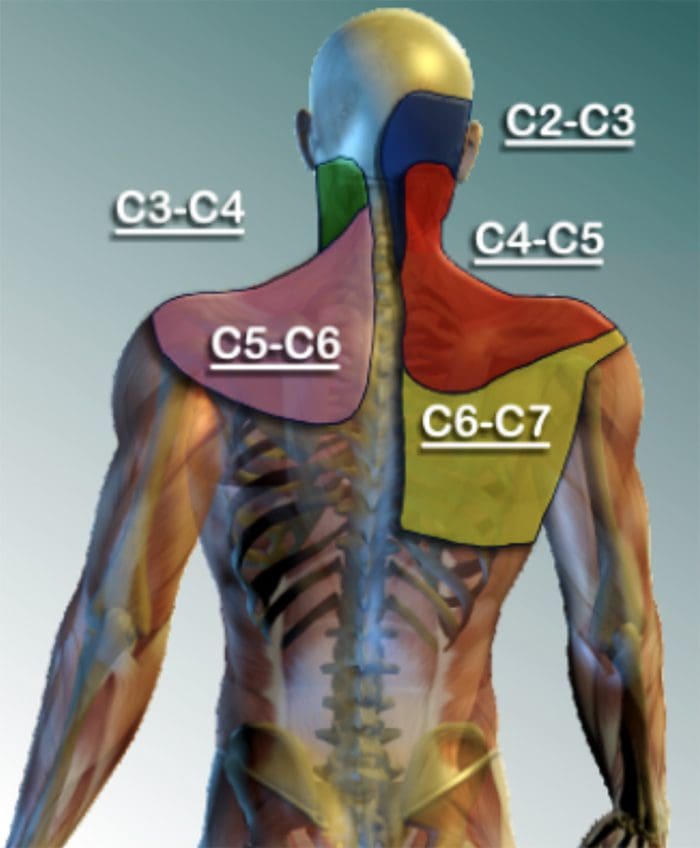
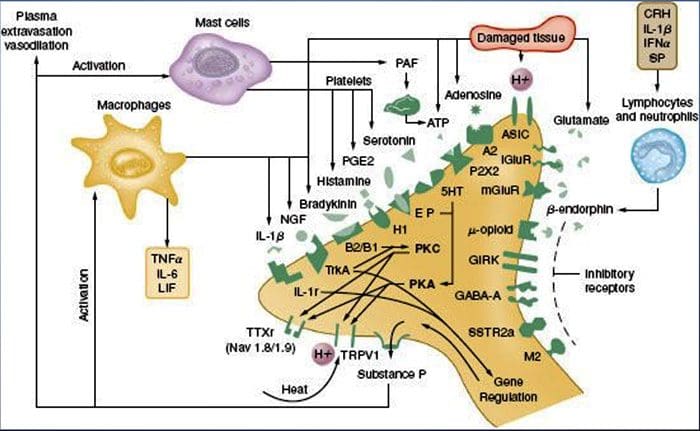 为什么我的肩膀会痛? 肩痛的神经解剖学和生化基础综述
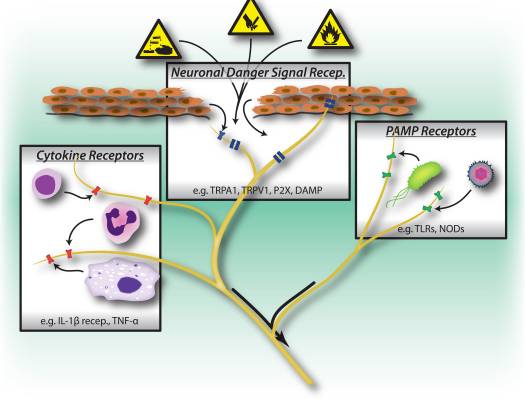
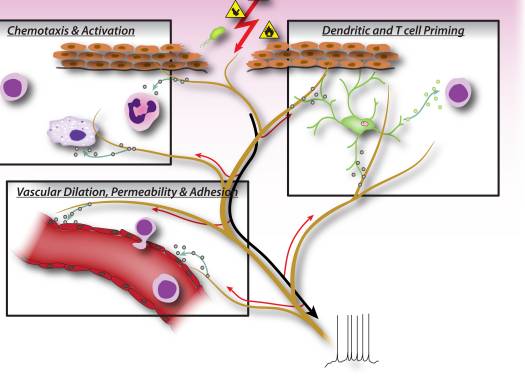
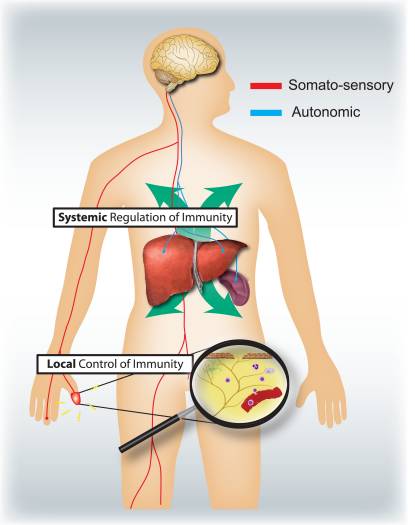
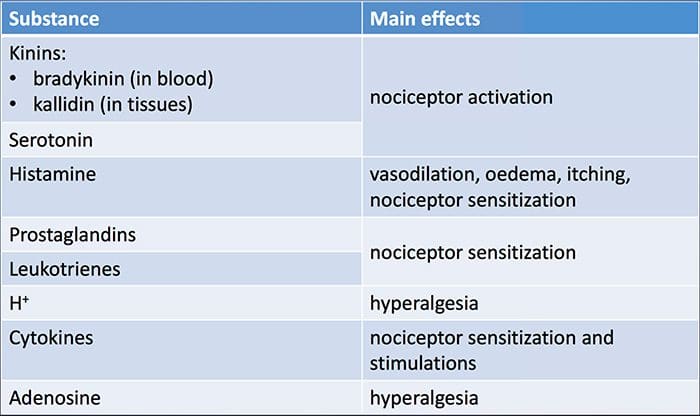
为什么我的肩膀会痛? 肩痛的神经解剖学和生化基础综述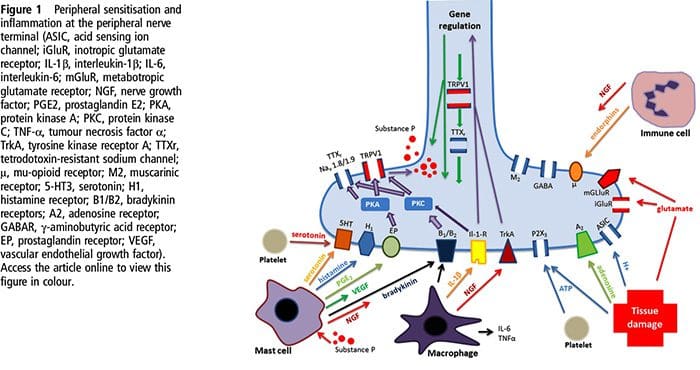 NGF 和瞬时受体电位阳离子通道亚家族 V 成员 1 (TRPV1) 受体在炎症和伤害感受器致敏方面具有共生关系。 发炎组织中产生的细胞因子导致 NGF 产生增加。 19 NGF 刺激肥大细胞释放组胺和血清素 (5-HT3),也使伤害感受器敏感,可能改变 A? 纤维使得更大的比例变得伤害性。 TRPV1 受体存在于初级传入纤维的亚群中,并被辣椒素、热量和质子激活。 TRPV1 受体在传入纤维的细胞体中合成,并被运输到外周和中央终端,在那里它有助于伤害性传入神经的敏感性。 炎症导致 NGF 在外周产生,然后与伤害感受器末端的 1 型酪氨酸激酶受体结合,然后 NGF 被转运到细胞体,导致 TRPV1 转录的上调,从而增加伤害感受器的敏感性。 19 20 NGF 和其他炎症介质也通过多种次级信使途径使 TRPV1 敏感。 许多其他受体,包括胆碱能受体、α-氨基丁酸 (GABA) 受体和生长抑素受体也被认为与外周伤害感受器敏感性有关。
NGF 和瞬时受体电位阳离子通道亚家族 V 成员 1 (TRPV1) 受体在炎症和伤害感受器致敏方面具有共生关系。 发炎组织中产生的细胞因子导致 NGF 产生增加。 19 NGF 刺激肥大细胞释放组胺和血清素 (5-HT3),也使伤害感受器敏感,可能改变 A? 纤维使得更大的比例变得伤害性。 TRPV1 受体存在于初级传入纤维的亚群中,并被辣椒素、热量和质子激活。 TRPV1 受体在传入纤维的细胞体中合成,并被运输到外周和中央终端,在那里它有助于伤害性传入神经的敏感性。 炎症导致 NGF 在外周产生,然后与伤害感受器末端的 1 型酪氨酸激酶受体结合,然后 NGF 被转运到细胞体,导致 TRPV1 转录的上调,从而增加伤害感受器的敏感性。 19 20 NGF 和其他炎症介质也通过多种次级信使途径使 TRPV1 敏感。 许多其他受体,包括胆碱能受体、α-氨基丁酸 (GABA) 受体和生长抑素受体也被认为与外周伤害感受器敏感性有关。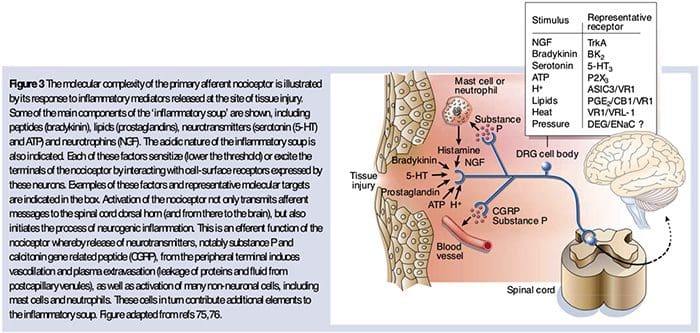 伤害感受器的神经化学
伤害感受器的神经化学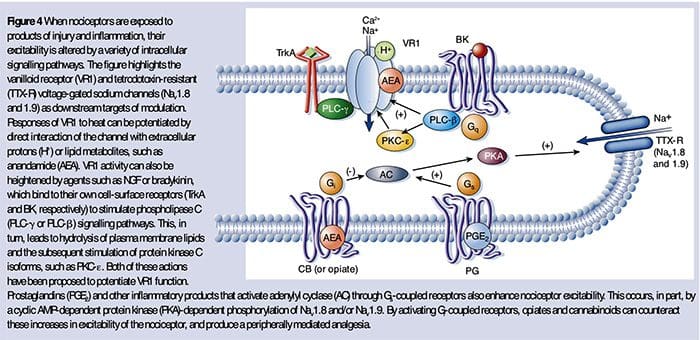 疼痛的细胞和分子机制
疼痛的细胞和分子机制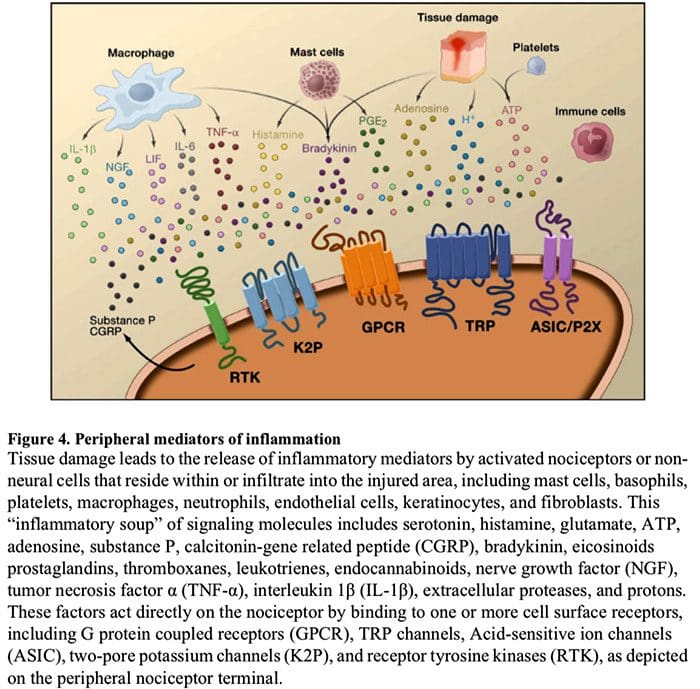
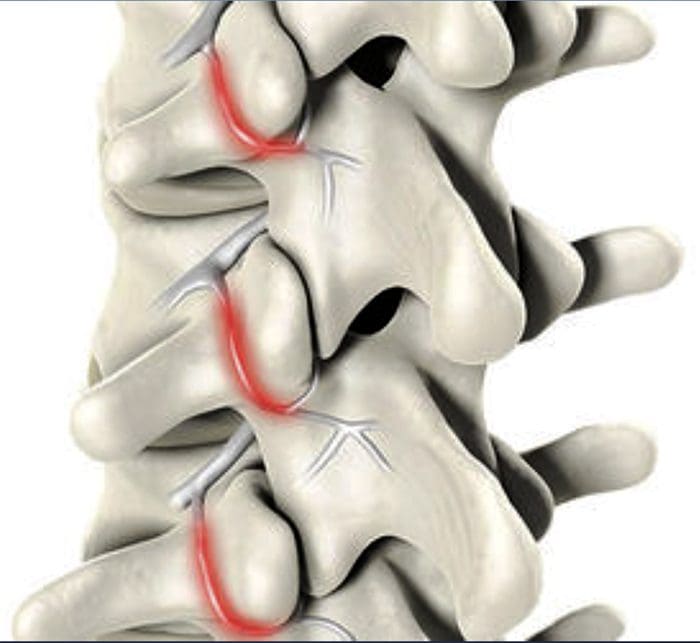
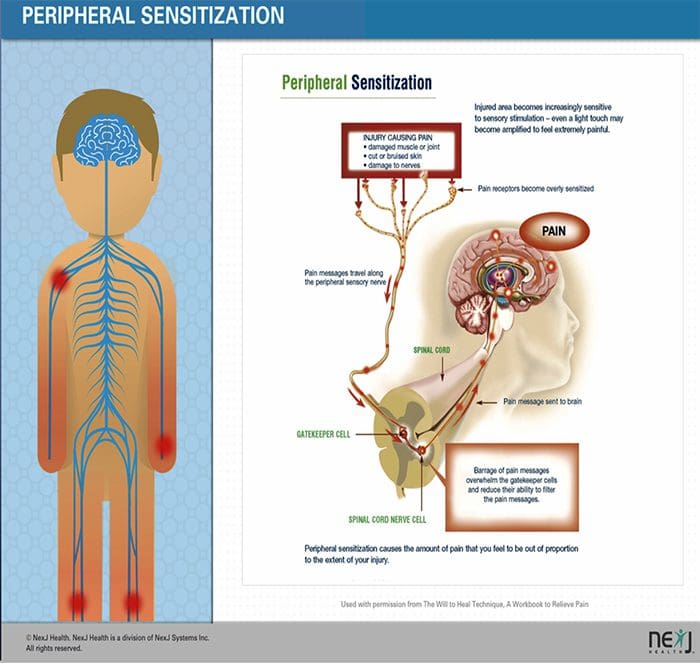
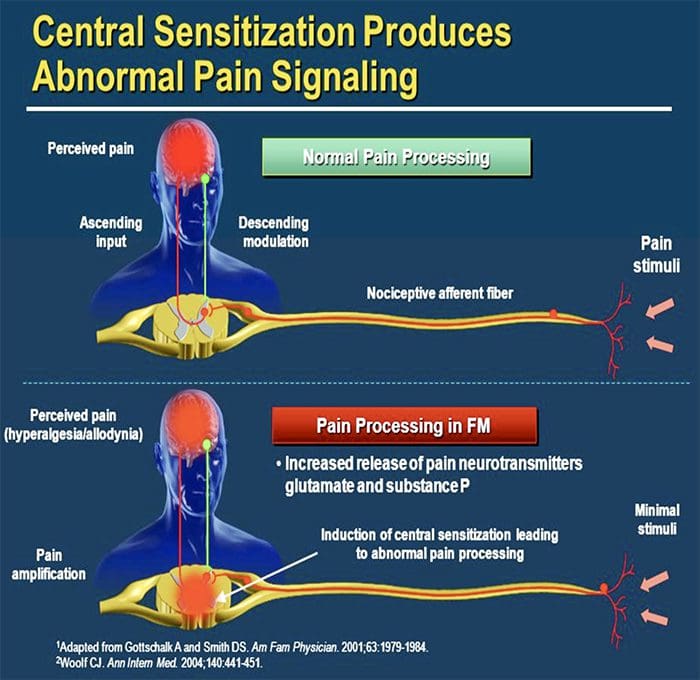
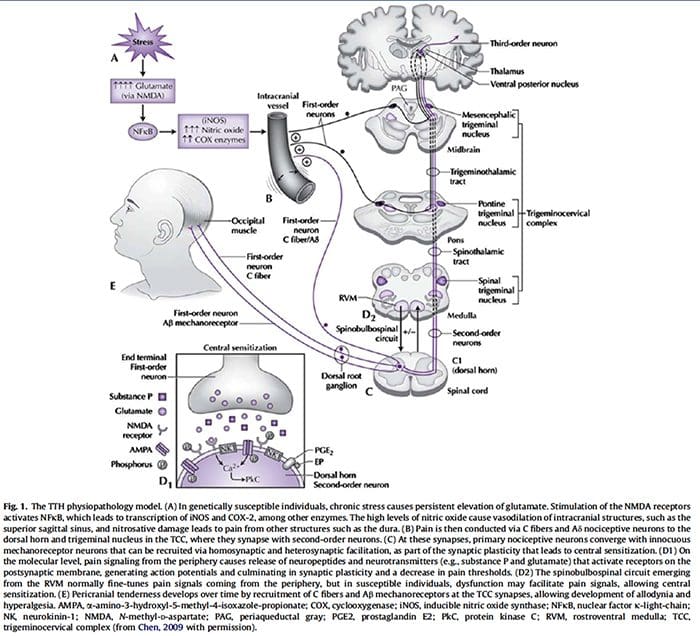
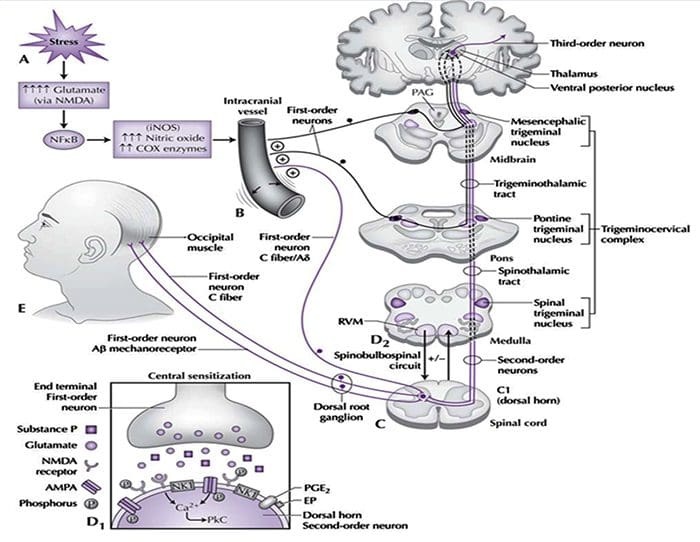
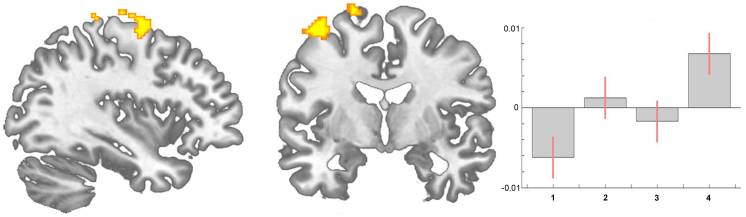
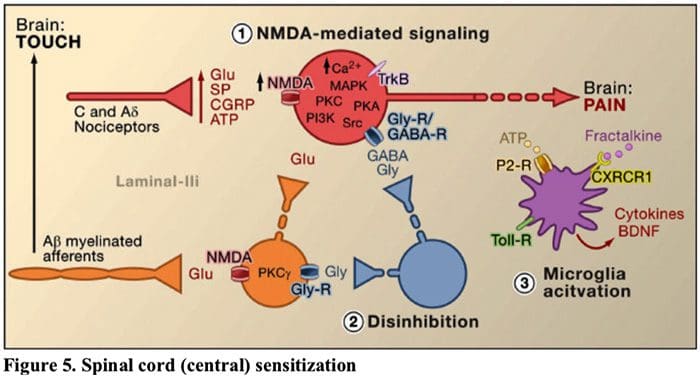 图 5. 脊髓(中央)致敏
图 5. 脊髓(中央)致敏